| Home | Syllabus | Schedule | Lecture Notes | Extras | Glossary |
| Home | Syllabus | Schedule | Lecture Notes | Extras | Glossary |
We began our introduction to prokaryotic genetics by pointing out that we had been using bacteria and their viruses as experimental organisms to investigate the biochemistry of heredity in prior lectures. Specifically, we investigated the nature of the genetic material with bacteria (Avery, MacLeod, and McCarty 1944) and with phage (the Hershey-Chase experiment, 1952). We investigated the nature of DNA replication with bacteria (the Meselson-Stahl experiment, 1958), and discovered mRNA using bacteria and phage (Brenner, Jacob, and Meselson 1961). Bacteria (mostly E. coli) and phage are convenient, fast, simple systems that allowed us to discover much of the biochemistry of the hereditary materail.
Why should we be interested in prokaryotic genetics? First, as we have already reviewed, these simple systems made it possible to make key advances in the science of genetics because of their simplicity. Second, humanity is always fighting bacterial and viral pathogens, so perhaps if we understood the genetics of E. coli and its phage, we could learn something that would help us in the fight against bacterial and viral pathogens. Third, these simple systems can be easily modified to become tools for molecular biology, genetic engineering, biotechnology, synthetic biology or whatever else we might call the practice of modifying living things as a kind of extension of chemical engineering.
Bacteria, especially E. coli, are easily grown in culture in liquid medium or on plates, as illustrated below. A suspension of cells is spread on an agar plate using a flame-sterilized spreading tool. If the plate is not selective (that is, it allows all of the bacteria to grow), we want to make sure that the suspension is sufficiently dilute to allow only a limited number of cells to be spread on the plate. Everywhere that a bacterial cell has landed will grow into a colony visible to the naked eye, with all cells in that colony being descendants of a single cell that landed on the plate. If the plate is selective (that is, contains an antibiotic to which the starting strain was not resistant), we can spread a very large number of cells to allow only rare resistant cells to form colonies.
Any study of genetics requires variants. Just as Mendel began the science of genetics with peas that varied for single traits, we need specific bacterial traits and variants that differ with respect to those traits. There are several kinds of variants of bacteria. We have already discussed auxotrophs, variants that require specific nutrients not normally required by that organism, for example the arginine auxotrophs in Neurospora that we discussed in our lecture on gene function. We can also study variants that are not able to use a specific energy source, for example the sugar lactose. We can also study variants that are resistant to things that normally kill bacteria, like antibiotics or phage.
Strangely enough, before bacterial genetics could get off to a good start in the 1940s, there was disagreement about whether bacteria even had heredity in the sense that people, pea plants, and Drosophila have heredity.
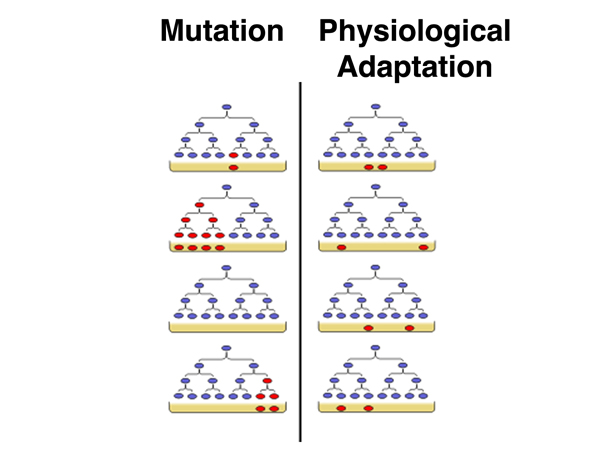
As difficult as it is for us to imagine this now, in the 1940s there were some who argued that bacteria did not have heredity in the same way that higher organisms do. Presented with evidence for "mutation" to antibiotic resistance in bacteria, some scientists argued that bacteria resistant to antibiotics were in an altered physiological state that is maintained among daughter cells when they divide. The frequency of entering this altered state was low, but it occurred when bacteria we forced to learn to adapt to the presence of an antibiotic.
The alternative hypothesis is that bacteria can mutate to antibiotic resistance, and that in any culture of bacteria there are some mutants whose presence is revealed upon exposure to the antibiotic. The key to testing these two hypotheses is that in the first case, that of physiological adaptation, the bacteria only shift to antibiotic resistance at a characteristic frequency upon exposure to the antibiotic. In the second case, the mutants are present before exposure to the antibiotic. How can we distinguish these possiblities?
While contemplating this problem, physicist Salvador Luria was attending a dance for scientists at which he observed another scientist feeding dimes to a slot machine. Luria explained to the gambler that his behavior made no sense, because each pull of the handle was an independent trial with negative expectations (some casinos even brag about a 98% payout on slots, giving the mathematically adept the knowledge that each pull of the handle on a $1 slot machine costs the player 2 cents on average). The slot player countered to Luria that playing slots was still a good game because, "You hit jackpots."
Luria immediately understood the implications of this statement. If a group of people play poker, the winners go home with money taken from the losers, but it is a closed system. In slots, an individual player may utterly defy the odds on a good day by winning a lot of other people's money on a jackpot on his first few plays.
Luria went straight to the lab an worked out an experiment to test which hypothesis was correct. In the image shown below, the predictions for the two hypotheses are shown. On the left, under the mutation hypothesis, cultures will differ in the overall frequency of resistant bacteria depending on whether mutations occurred early in the growth of the culture (a jackpot) or late. On the right, under the hypothesis of physiological adaptation, because the bacteria do not adapt until challenged, the frequency of resistant bacteria will be more or less constant in any sample.
Luria and Delbruck used resistance to bacteriophage T1 as the characteristic to study. They grew one culture in a flask for a while before subdividing it into twenty small cultures that they grew for an additional time. They grew a second culture in a flask without subdividing it. At the end of the growth period, they plated bacteria from the subcultures or the single culture on plates that they sprayed with bacteriophage T1. The results are shown below in cartoon form.
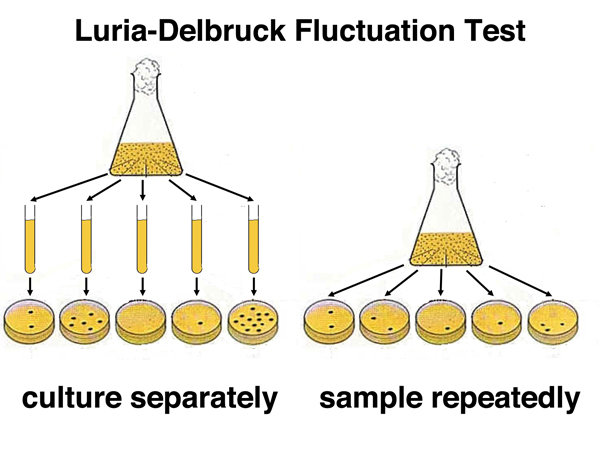
The plates from the subcultures showed a wide range in the frequency of T1-resistant cells in each subculture. Some lucky subcultures hit the jackpot and had a high frequency of resistant cells (mutants arose early in the growth of the culture). The plates from the combined culture had more or less the same frequency of resistant cells on each plate. This showed that bacterial variants arose by mutation in advance, rather than by physiological adaptation upon a challenge.
This experiment was presented in a paper that analyzed the results with considerably more mathematical sophistication than is used here. Consequently, it was not widely accessible to all biologists, but fortunately another type of experiment was able to establish the pre-existence of bacterial mutations to the average mathematically-challenged biologist.
Lederberg and Lederberg invented replica plating. In this technique, a sterile velevet is pressed against a culture plate on which bacteria have grown, then pressed against a series of sterile agar plates to make "copies" of the orginal plate, as shown below.
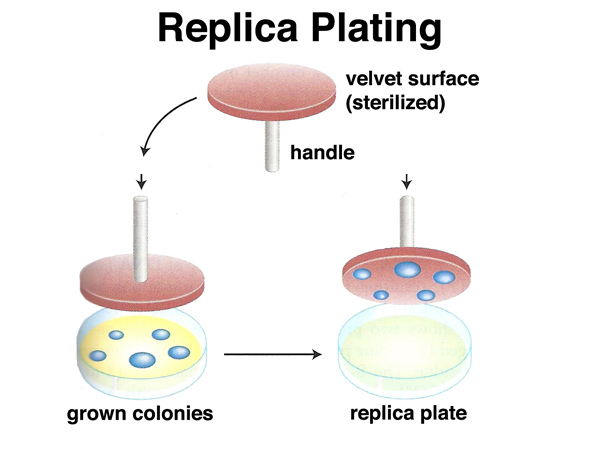
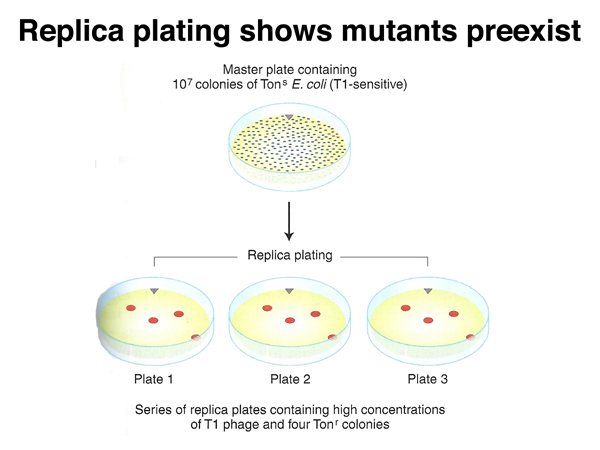
A very dense plate of 107 colonies of E. coli that is sensitive to infection by phage T1 can be used as the master plate to make a series of replica plates. Each of the replica plates is then sprayed with T1 and incubated, as shown below. If T1-resistant mutants exist on the master plate, they will appear on the replicas in the same pattern. If T1 resistance is only induced upon exposure to phage, the resistant colonies that appear on the replica plates will appear in different locations on each plate.
Now that we have established that we can identify genetic variants of bacteria, we need to find out if there is any way to mate them with each other. More generally, we need to know if there is any mechanism to transfer genetic information from one bacterial cell to another.
There are three mechanisms by which DNA can move from one bacterial cell to another:
These three mechanisms are outlined in the figure below.
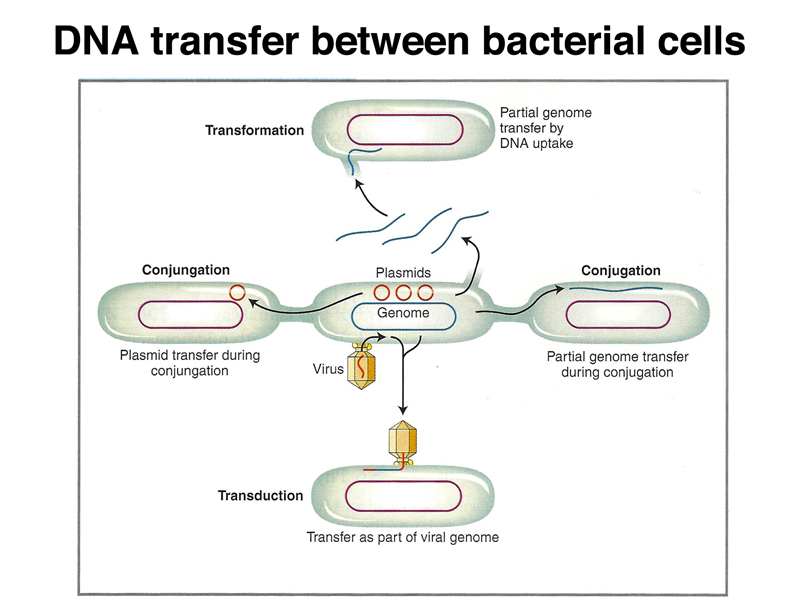
Transformation. We will not spend any additional time discussing transformation.
Conjugation. Lederberg and Tatum discovered the process of bacterial mating in 1946. They mixed two different strains with two or three auxotrophic mutations each, then plated the mixture on minimal medium. While neither strain was capable of growing on minimal medium by itself, among the bacteria grown in mixed culture were fully prototrophic cells capable of growning on minimal medium, as shown below. This suggests that bacteria are capable of mating.
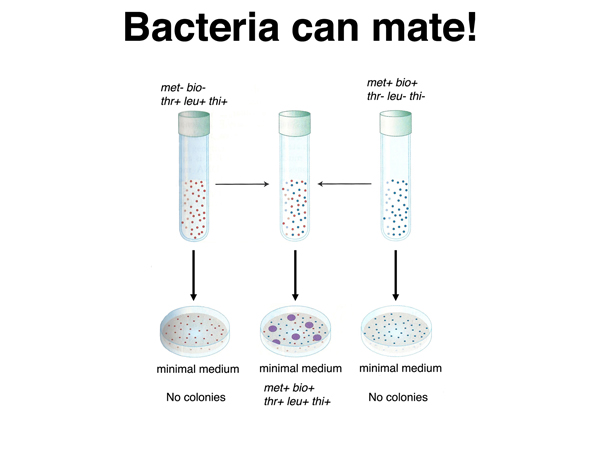
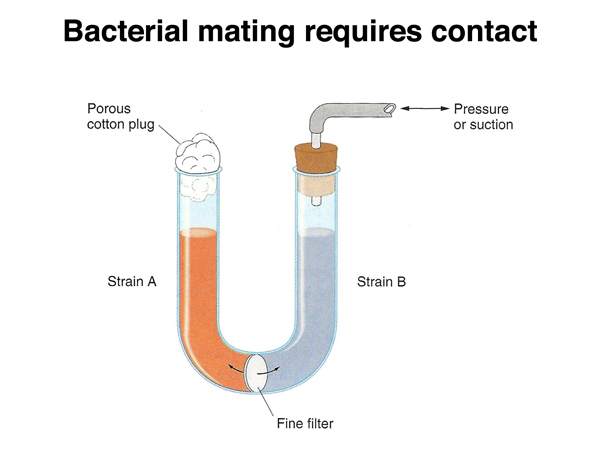
The bacterial mating process requires contact between cells. In the experiment shown in the figure below, the two strains were cultured in a U-tube, with air pressure used to move medium between the two parts of the tube through a filter. When the pores of the filter are too small to allow bacteria to pass through them, no prototrophic cells are recovered.
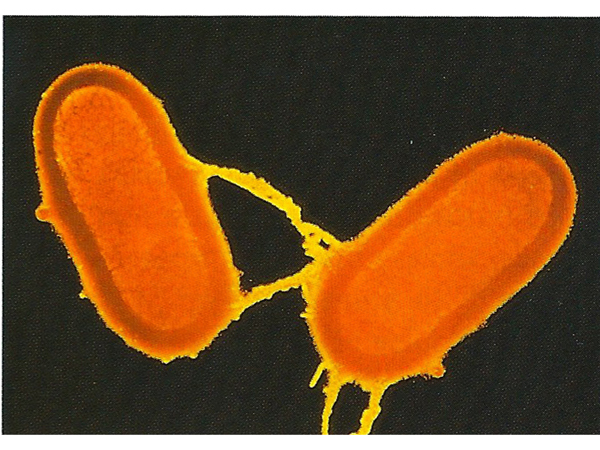
The image below shows the appearance of bacterial cells during this mating process, called conjugation. Conjugation in bacteria is unidirectional. A donor strain makes contact with a recipient strain using a tube called a pilus.
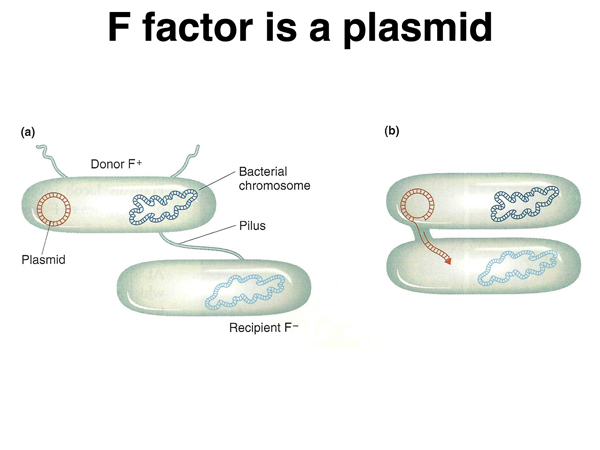
It was discovered early in the study of conjugation that fertility can be lost. The fertility of a strain that has lost fertility can be restored by mixing it with a fertile strain. The fertility factor, or F factor, can be thought of as infectious. The F factor is a plasmid, a small piece of circular DNA that is separate from the bacterial chromosome. The F factor causes strains harboring it to have pili, and to transfer the F factor to recipient strains, as shown below.
Transfer of the F factor occurs by rolling circle replication, as shown below. One strand is nicked. DNA replication from the 5' to 3' direction displaces a free 5' end, which is transferred through the pilus to the recipient strain. As single-stranded DNA enters the recipient strand the other strand is synthesized.
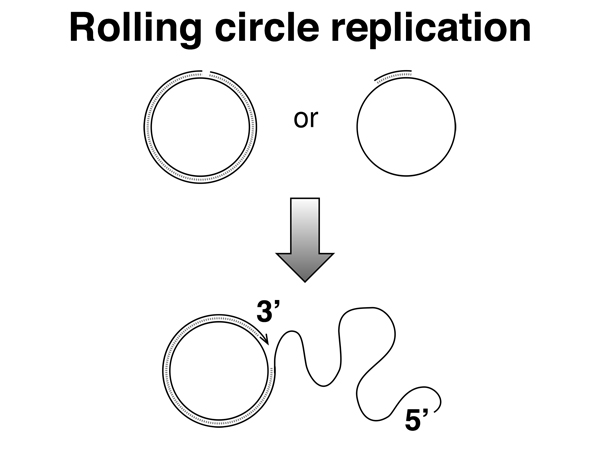
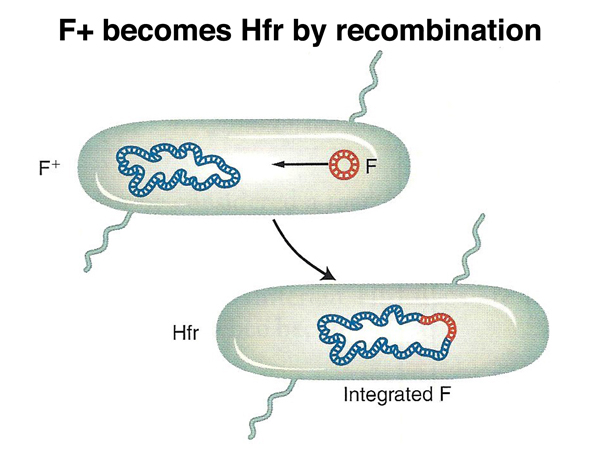
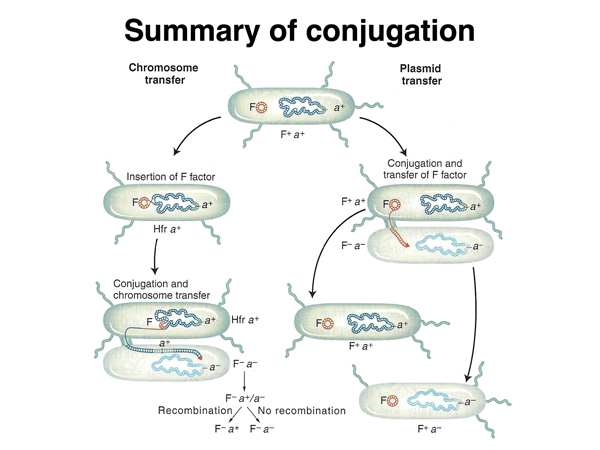
Transfer of the F factor occurs at much higher frequencies than the transfer of chromosomal DNA from the donor strain. An unusual strain was recovered that transferred chromosomal DNA at 1000x the frequency of other F+ strains. This strain was named "high frequency recombination" or Hfr. Besides transferring chromosomal DNA at a much higher frequency, this strain rarely transferred fertility to the recipient strain. In the Hfr strain, the F factor had recombined into the bacterial chromosome, as shown below.
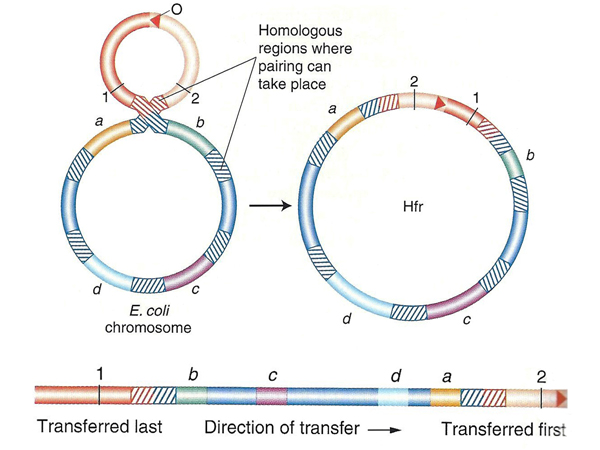
As shown below, there are multiple sites of homology to the F factor in the E. coli chromosome. Recombination between the F factor and any of these sites will create an Hfr strain. Transfer of a copy of the donor chromosome to the recipient strain occurs using the same mechanism that is used to transfer the F factor. The F factor sequence is nicked to initiate rolling circle replication. Because part of the F factor is transferred last, after the entire bacterial chromosome has been transferred, most matings do not succeed in transferring fertility to the recipient strain.
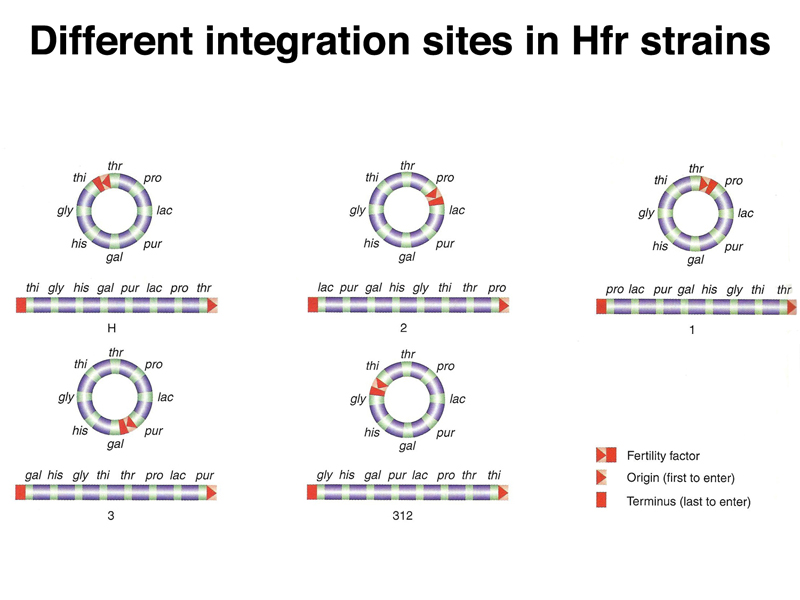
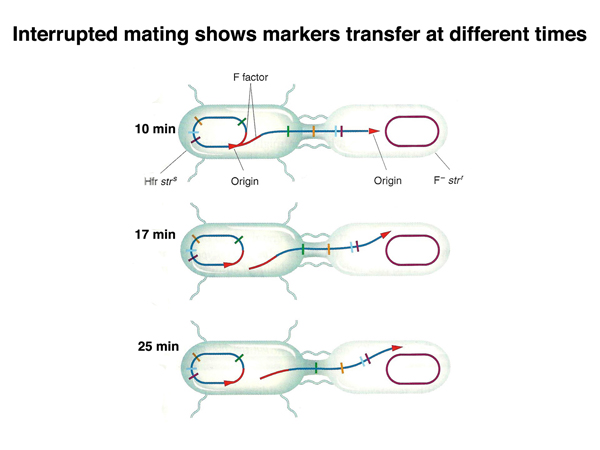
The use of different integration sites will create different Hfr strains that initiate transfer at different locations in the E. coli chromosome, as shown below. Each integration site will produce a particular direction of transfer, clockwise or counterclockwise on the standard E. coli chromosome map.
As shown below, as the chromosome is transferred to the recipient strain, the other strand is synthesized. In order for any of the donor markers to be inherited by the recipient, recombination must take place. When a linear molecule recombines with a circle, all donor markers between two crossovers will replace the corresponding alleles in the recipient.
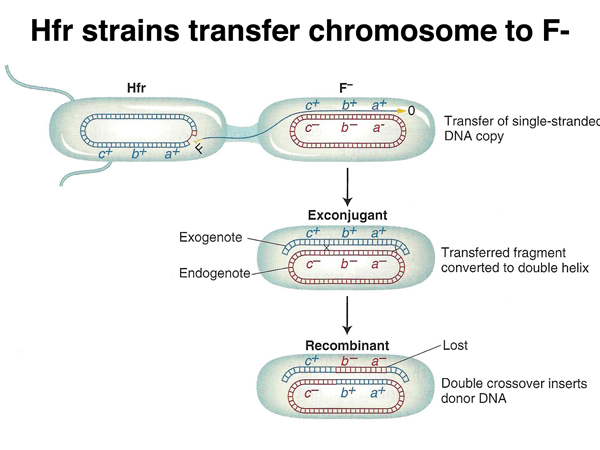
What is the mechanism by which F+ strains transfer chromosomal DNA to recipients? It turns out that this occurs by the formation of Hfr strains when the F+ strain is grown in culture. This is summarized below. If there is no recombination between the F factor and the chromosome, only the F factor can be transferred.
Any particular Hfr strain always transfers markers in the same order. This allows us to use time of transfer as a way of constructing genetic maps. We can interrupt mating at various times after mixing the cultures by blending the mixed culture in a blender before plating on selective medium. In the experiments summarized below, the order of transfer of markers is azi - ton - lac - gal.
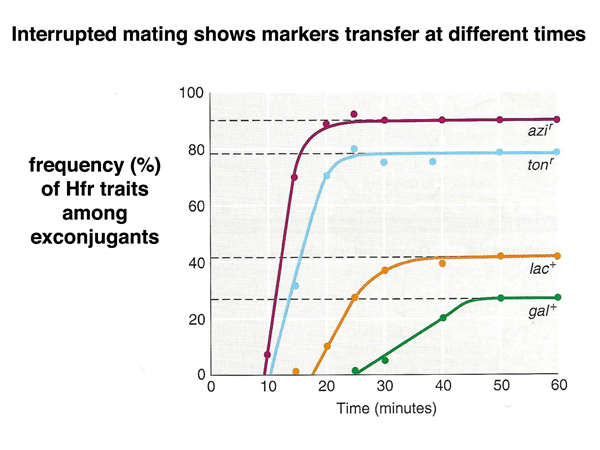
The figure below shows mapping by time of marker entry in an Hfr x F- cross in cartoon form.
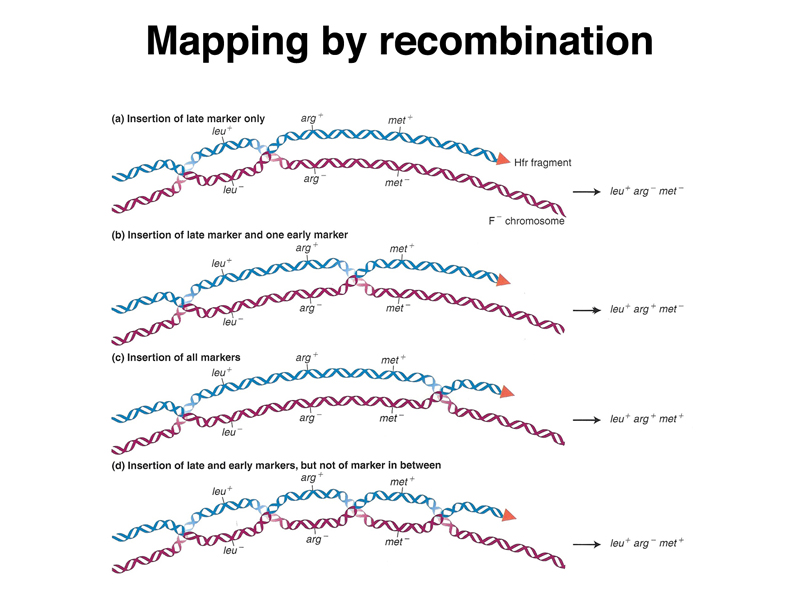
The crossovers that allow the incorporation of donor DNA into the recipient chromosome can occur in different places to produce different outcomes. The figure below shows (a) insertion of a late maker only as a consequence of a pair of crossovers flanking that marker, (b) insertion of a pair of markers as a consequence of crossovers flanking those markers, (c) insertion of three markers by widely separated crossovers, and (d) insertion of two markers, but not the marker between them, as a consequence of three crossovers. Transfer of markers from an Hfr strain can be used to map the order of markers and the relative distance between them in a way similar to linkage analysis in eukaryotes.
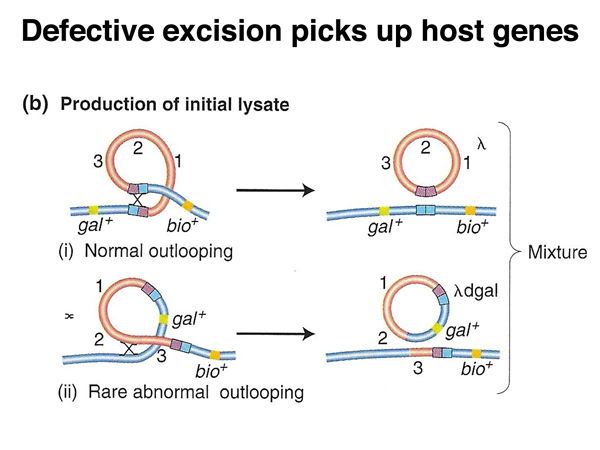
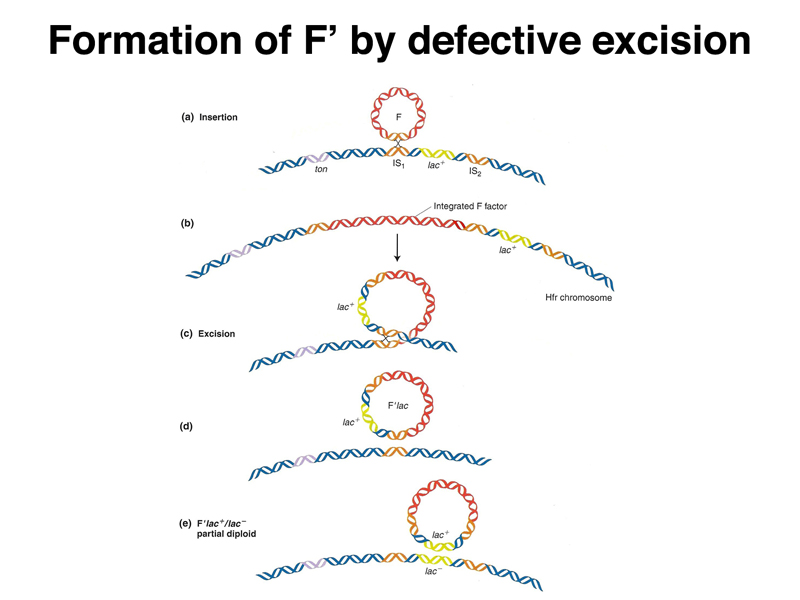
As shown below, it is possible for the F factor to excise itself from the host chromosome in an Hfr strain. At a low frequency, the recombination events will occur in different sequences than the sequences used for integration. In such a defective excision, the F factor will pick up a region of the bacterial chromosome. An F factor carrying a portion of the host chromosome is called an F' (F prime). An F' strain will transfer the portion of the host chromosome carried on the plasmid at high frequency, and when the F' integrates into the chromosome, it is likely to do so at the site of homology to the host chromosome.
Multidrug resistance plasmids. No discussion of plasmids is complete without a brief mention of multidrug resistance plasmids. These were first discovered in the 1950s in Shigella, a bacterium that causes dysentery. A strain appeared that was resistant to several different antibiotics. The antibiotic resistance genes were all carried on a plasmid that is made up of sequences derived from several different bacterial species. The multidrug resistance plasmid can be easily transferred between species. This is one of the reasons that infections acquired in hospitals are so dangerous; multidrug resistance plasmids that can be transferred between species are common in hospitals.
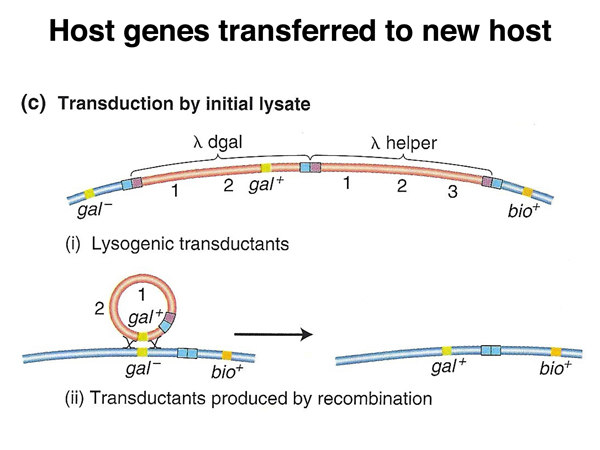
Transduction. Transduction is the transfer of bacterial DNA from one cell to another via a bacteriophage. There are two kinds of transduction: generalized transduction, in which any gene can be transferred, and specialized transduction, in which only certain genes are transferred. Any particular type of phage will usually only be capable of one type of transduction.
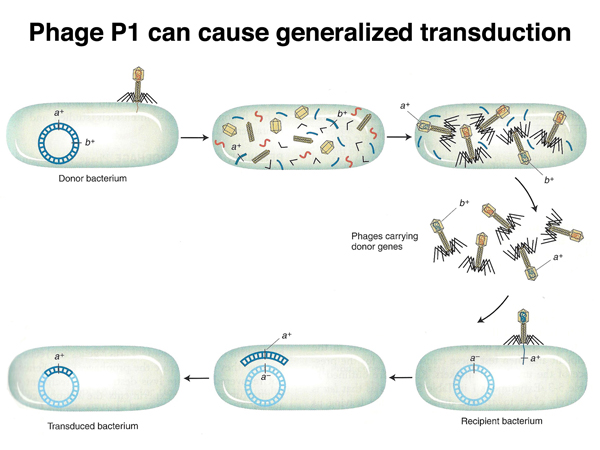
Phage P1 is a generalized transducing phage. P1 is only capable of producing a lytic infection, in which the bacterial cell lyses, releasing hundreds of phage particles. About 1/10,000 of the P1 phage particles carry chromosomal DNA rather than phage DNA. If a P1 lysate from a donor strain is used to infect a recipient strain at low multiplicity of infection (meaning that any cell of the recipient strain is not likley to be infected by more than a single phage particle), some of the recipient cells will receive chromosomal DNA from the donor cell. Recipient cells that have incorporated donor DNA can usually be selected.
Not all phage produce lytic infections all of the time. Phage λ (lambda) is a temperate phage that in addition to lytic infections can become a lysogen by integrating into the E. coli chromosome at a specific location, as shown in the figure below.
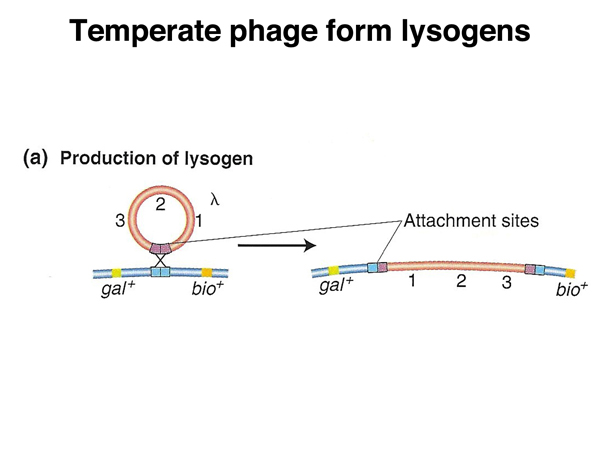
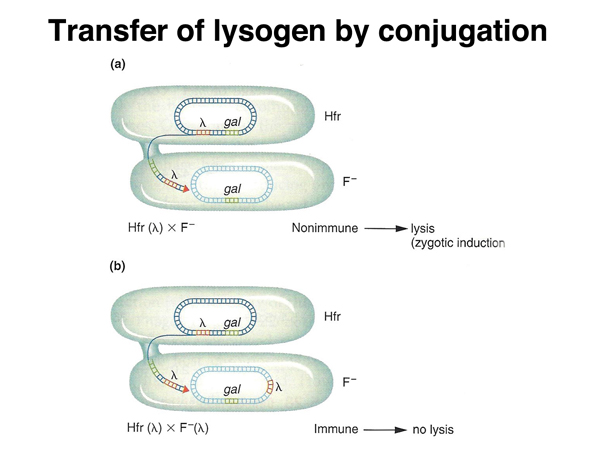
A λ genome integrated into the host chromosome is called a lysogen because it induces lysis under particular circumstances. If an Hfr strain that is a lambda lysogen is mated to an F- strain that is not a lamda lysogen, the phage is induced, and the recipient cell undergoes a lytic infection. If the F- cell is a lambda lysogen, induction of the phage does not occur. This tell us that in cells that are lysogens, there is a mechanism to keep the prophage (the integrated λ genome) from intiating a lytic infection. If the lambda genome is transferred to a non-lysogenic cell during mating, the cellular factor that keeps the prophage inactive (a protein) is absent, and a lytic infection is initiated, as shown below.
Phage λ is a specialized transducing phage. Defective excision of the λ genome will produce defective phage that can transduce the flanking gal or bio genes, as shown in the diagrams below.