
| Home | Syllabus | Schedule | Lecture Notes | Extras | Glossary |
| Home | Syllabus | Schedule | Lecture Notes | Extras | Glossary |
We began by reminding all students to stay on top of this course, checking the schedule to make sure that they know when reading quizzes and homework assignments are due. Most students are reading the lecture notes.
The deadline for a short essay (no more than 1,000 words) on what you might learn from having your genome analyzed is midnight on Friday, September 13. The best assay will win a free genome analysis from 23andMe. This is not a class assignment, and no extra credit will be awarded for participation. You will not be required to share any of the results of the analysis of your genome with anyone, ever. You can read about one person's experience at OpenHumanGenome.
We have also frequently repeated the advice that success in this course might require changing the way that you think. For today's material, changing the way that you think would mean focusing less on the answers that we deduce from the experiments, which after all might be modified by the results of further experiments, and more on the process of reasoning that is used in genetic analysis.
We reviewed the underpinnings of today's lecture, which seeks to unify Mendelism with the behavior of chromosomes. We know from Mendel's work that hereditary units are particles called genes that are not affected by their passage through hybrids. We know from the work of Flemming, Sutton, and Boveri that cells use an elaborate mechanism to ensure the accurate transmission of chromosomes to cells in both mitosis and meiosis. We reviewed Mendel's First Law, the principle of segregation, which says that the two alleles of a gene in a hybrid segregate into different gametes. We reviewed Mendel's Second Law, the principle of independent assortment, which says that the two alleles of a pair of genes in a dihybrid will segregate independently during the formation of gametes.We also saw that the Second Law is rooted in the behavior of different bivalents at metaphase of meiosis I. The two different bivalents have no way of communicating with each other, so the two different maternally-derived centromere regions are as likely to orient to the same pole of the spindle apparatus as they are to orient to different poles. This reminds us that chromosomes don't follow Mendel's rules, they make them.
Morgan's lab at Columbia University began studies of the fruit fly Drosophila melanogaster, which had many properties that made it an ideal organism for the study of genetics. Drosophila breed quickly, with a generation time of about 10 days. An individual female can lay several hundred eggs, so it is possible to accumulate a lot of genetic results quickly. Drosophila are also easy to grow and handle.
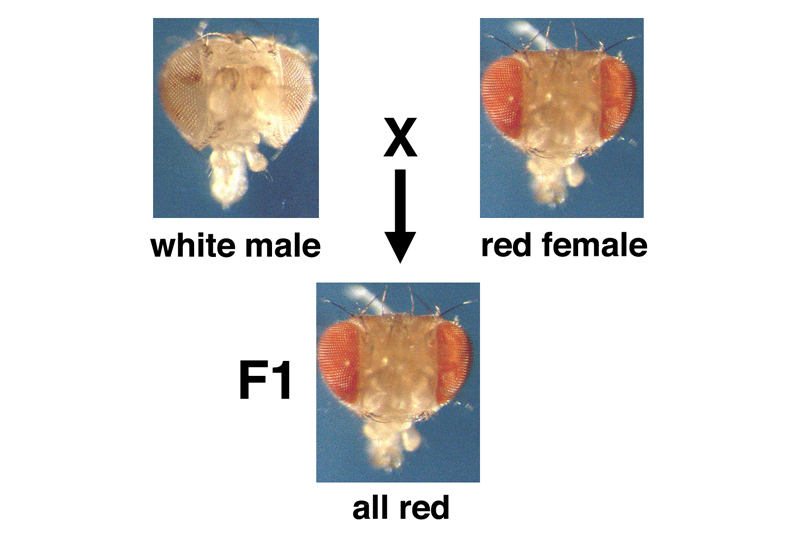
One day, a striking variant appeared that had white eyes instead of the usual brick red. The individual was a male, which was crossed to normal red-eyed females. The F1 progeny were all wild type (with red eyes), as shown in the picture below. This tells us that the variant is recessive.
An intercross of the F1 red-eyed males and females produces the familiar results shown below, displaying the expected 3:1 ratio seen from a monohybrid cross.
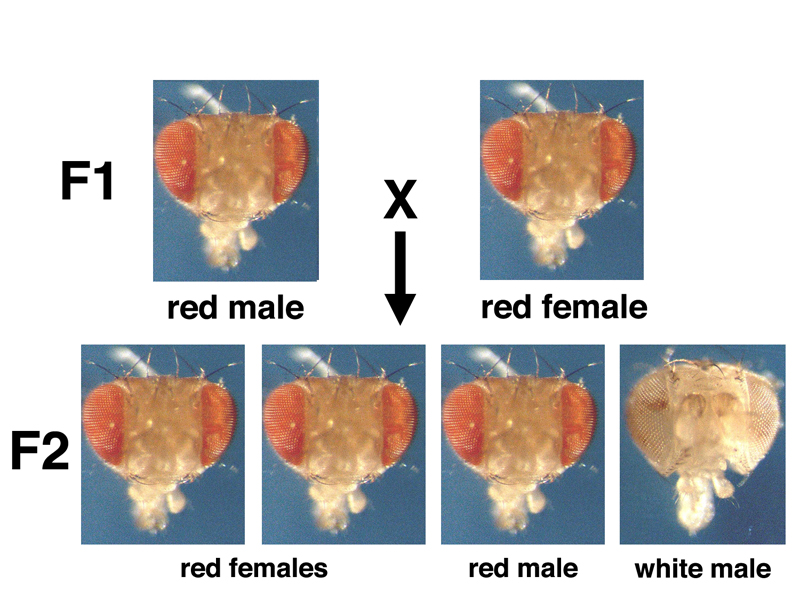
Yet these results are actually very peculiar. All of the white-eyed progeny are male. The phenotype of white eyes is linked to sex. It is also possible that white eyes is a sex-limited trait like pattern baldness in humans. Further experiments will resolve this.
We know that the 3:1 ratio in a dihybrid cross reflects a genotypic ratio of 1:2:1. This suggests that it is at least possible that some of the F2 females are hybrids rather than true-breeding wild type. Crossing F2 white-eyed males to F2 red-eyed females gives the results shown below for half of the females.
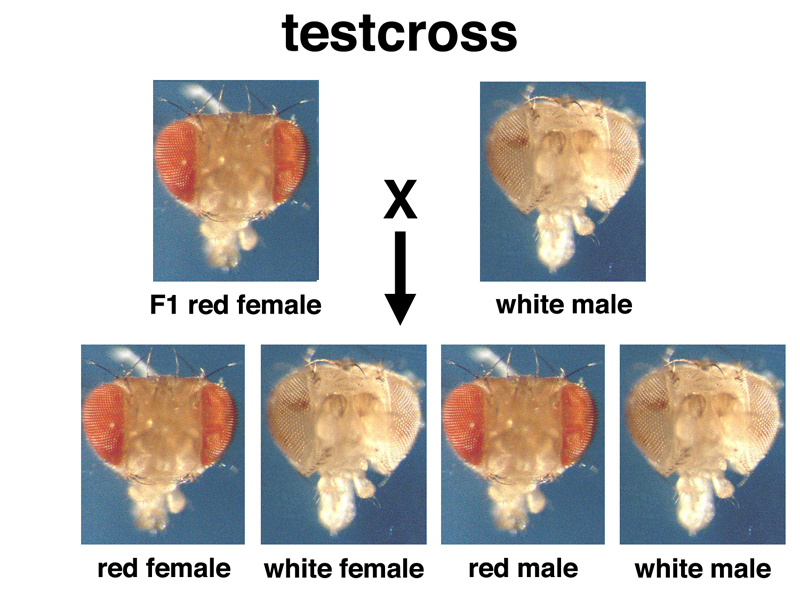
This 1:1:1:1 ratio of the four types withe respect to sex and eye color makes it evident that white eyes is not a sex-limited trait, and white-eyed females are possible. Crossing white-eyed males to white-eyed females gives a true-breeding line with white eyes, which makes further testcrosses possible.
When white-eyed males are crossed to red-eyed females, the F1 hybrids all have red eyes, as seen before. We can now cross red-eyed F1 males or females to white-eyed flies from our true-breeding line in a testcross. If we cross F1 red-eyed males to white-eyed females, we get the results shown below.
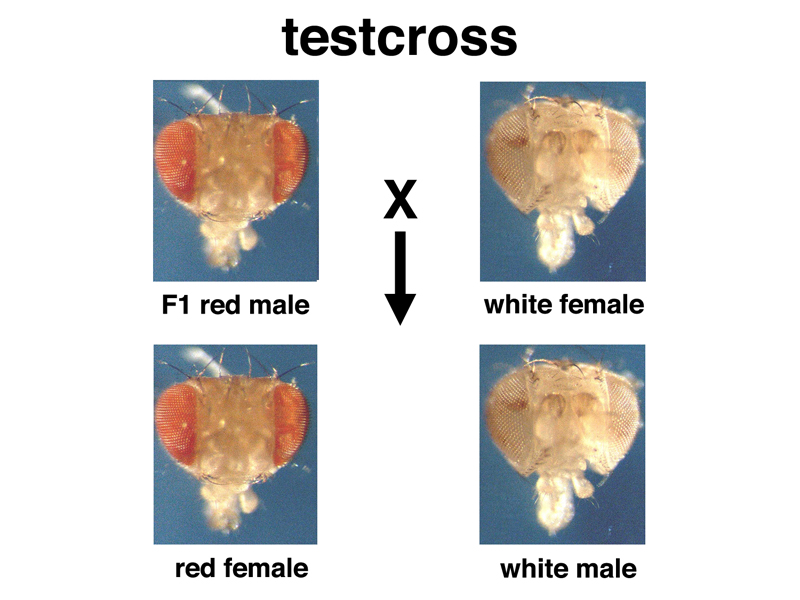
We get the expected 1:1 ratio of red eyes to white in the testcross of the F1 hybrids, but the results are very strange: all of the white-eyed progeny from the testcross are male. There aren't any red-eyed males. All of the red-eyed progeny from the cross are female. There aren't any white-eyed females.
When we cross F1 red-eyed females to white-eyed males, we get the results shown below.
In this testcross, we get the expected 1:1 ratio of red to white, but this time there are both red-eyed and white-eyed males and females in a 1:1:1:1 ratio.
What is different about these two crosses? Hybrid F1 males transmit the recessive allele only to their sons. Hybrid F1 females transmit the recessive allele to both sons and daughters.
What else to males transmit only to their sons? The Y chromosome.
This makes us think that the Y chromosome does not carry the genes for red eyes found on the X chromosome, and that perhaps that the Y chromosome does not carry many of the genes found on the X chromosome. Even at the time of these experiments, it was known that there are a number of human genetic diseases that are much more common in males than in females, for example hemophilia and color blindness.
We can use Punnett squares to evaluate the hybothesis that the white gene is on the Drosophila X chromosome. This hypothesis will make specific predictions.
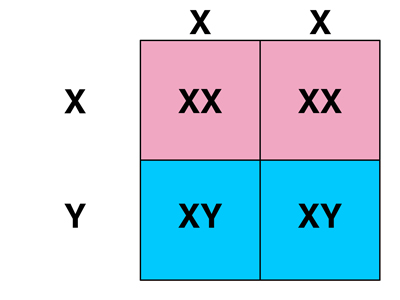
First, let's consider the cross of white-eyed females to red-eyed males (from the F1 or from a true-breeding red-eyed stock, it shouldn't matter). As shown in the frist Punnett square below, the sex of an individual will depend on which chromosome was inherited from the male parent: eggs that are fertilized by X-bearing sperm will be XX females, while eggs that are fertilized by Y-bearing sperm wil be XY males.
If we hypothesize that the white gene is on the X chromosome and that this gene is not found on the Y chromosome, we can fill in the genotypes and phenotypes as shown in the second Punnett square below.
 |
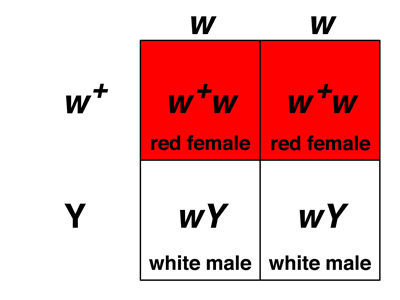 |
Our hypothesis predicts that the cross of white-eyed females to red-eyed males should produce a 1:1 ratio of red-eyed females to white-eyed males, which is exactly what we observe experimentally.
What about the other cross, of F1 hybrid females to white-eyed males in a testcross? The rules for chromosomal sex determination are the same, as shown in the first Punnett square below, but in this cross the hybrid females make two kinds of eggs: those that carry an X chromosome bearing the recessive w allele, and those that carry an X chromosome with the dominant w+ allele. Because both males and females inherit an X chromosome from their mothers, the two kinds of eggs produce either red-eyed or white-eyed females (if fertilized by a X-bearing sperm with the w allele) or red-eyed or white-eyed males (if fertilized by a Y-bearing sperm), as shown in the second Punnett square below.
|
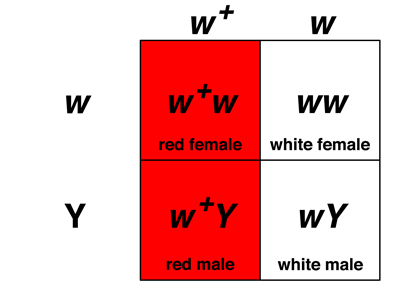 |
Our hypothesis predicts that the cross of red-eyed F1 hybrid females to white-eyed males should produce a 1:1:1:1 ratio of red-eyed and white-eyed individuals of both sexes, which is exactly what we observe experimentally.
We conclude, at this time, from our experiments with white:
We will return to the consideration of Drosophila experiments after a brief exploration of human pedigrees.
Our consideration of sex-linked inheritance in humans begins with a very famous individual, Queen Victoria of England (1819 - 1901), the longest-reigning monarch in British history. Queen Victoria is known to have been a carrier of a recessive variant allele of the sex-linked gene for coagulation factor IX (F9). This particular variant allele is responsible for the inherited disease hemophilia, in which blood clotting is defective.
We looked at a pedigree of Queen Victoria's many descendants. The line of succession to the British monarch went through Edward VII, then George V, then George VI, the father of the current British monarch, Queen Elizabeth. Edward VII was not a hemophiliac, so the current British royal family is free of the disease. One of Queen Victoria's other sons, Leopold, was a hemophiliac, whose daughter Alice was a carrier (as are all daughters of males with the variant). Alice had a hemophiliac son, Viscount Trematon, who died at age 20. The pedigree shows ten affected males in total, many of who died young, and no affected females.
We can identify carrier females either because their father was a hemophiliac (all such daughters are carriers), or because they gave birth to sons with hemophilia.
The typical appearance of human pedigrees in which a rare sex-linked variant is segregating is reduced to its essence below. Carrier females marrying normal males are both healthy, but half of their sons are affected. Half of their daughters are carriers.
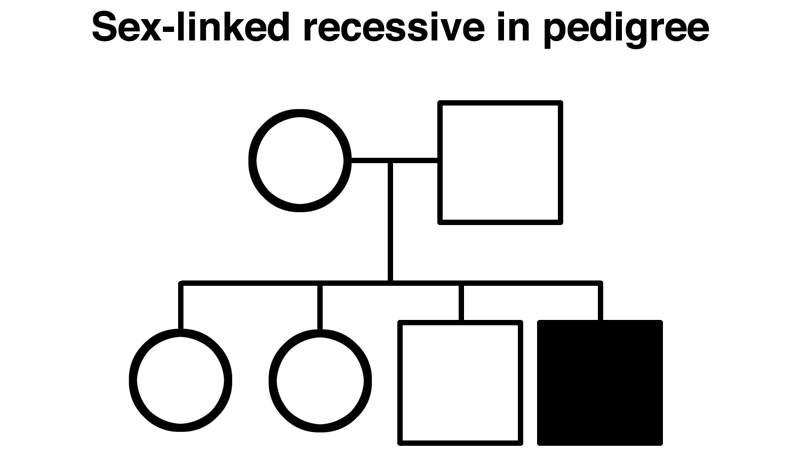
This result looks exactly like the F1 intercross with the white gene of Drosophila, shown below.
We continued our discussion of pedigrees with tips on how to solve them quickly, adapted from a video by Andrew Douch, posted on the Extras page. When we first begin to analyze pedigrees, we are really evaluating four different hypotheses. A variant allele can be either dominant or recessive, and it can be either autosomal or sex-linked. In general, we cannot prove a hypothesis in science, but we can disprove one. Solving pedigrees means eliminating possibilities until we are left with only one.
There are a variety of things that could make pedigrees harder to solve: false paternity, de novo mutation to give rise to new variants, incomplete penetrance, and so on, but for most purposes the approach outlined here will work well and it is fast, a real advantage on tests.
Consider the pedigree shown below.
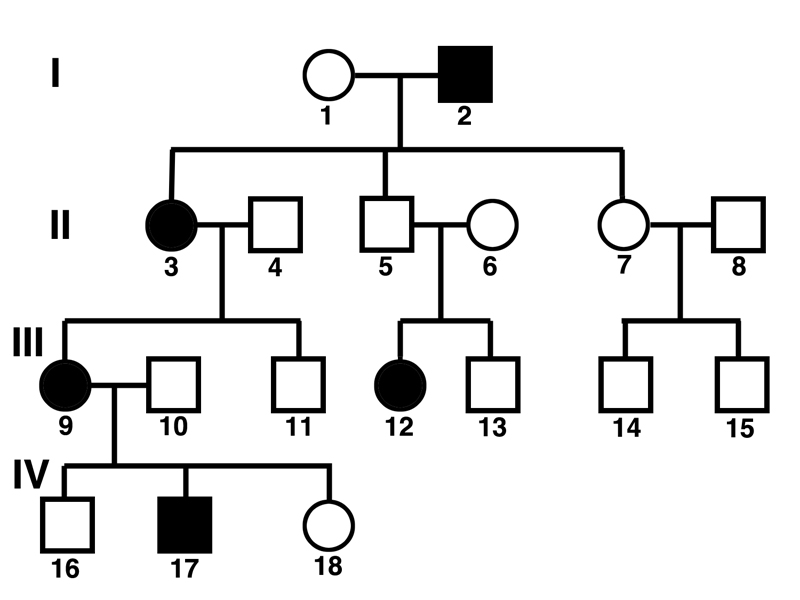
Andrew Douch points out that we should ask ourselves two questions:
Taking on the first question, we can see that individual 12 is an affected female whose mother (6) and father (5) are both unaffected.
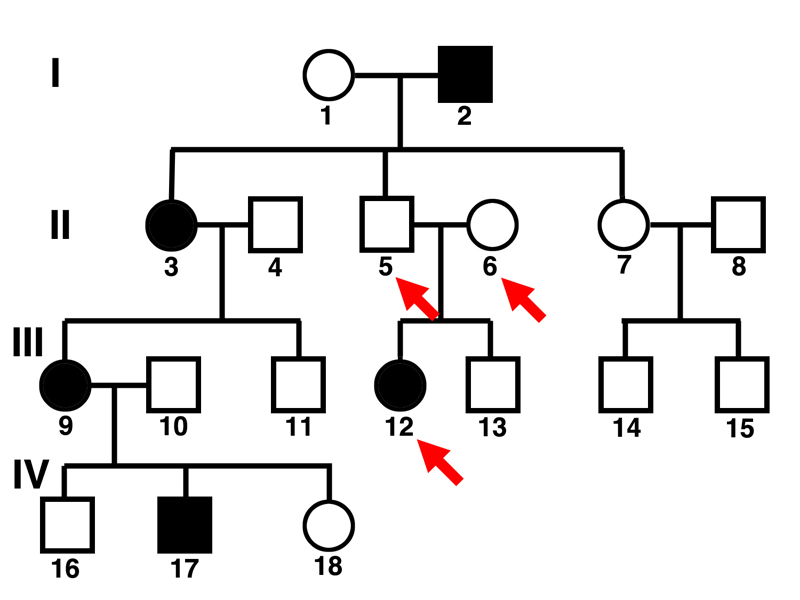
This tells us that we are dealing with a recessive trait. Two of the hypotheses (autosomal dominant and sex-linked dominant) are eliminated.
Taking on the second question, we can see that there are several examples of affected females who have fathers or sons that are unaffected. If an affected female has an unaffected father, the gene cannot be sex-linked, because women get one of their X chromosomes from their fathers, so if the father has an X chromosome that has the normal allele, the woman will be at least heterozygous, and cannot be affected by a sex-linked recessive allele. All of an affected woman's sons will be affected if the gene is a sex-linked recessive, because all sons get their X chromosomes from their mother.
The second question is addressed in the pedigree below.
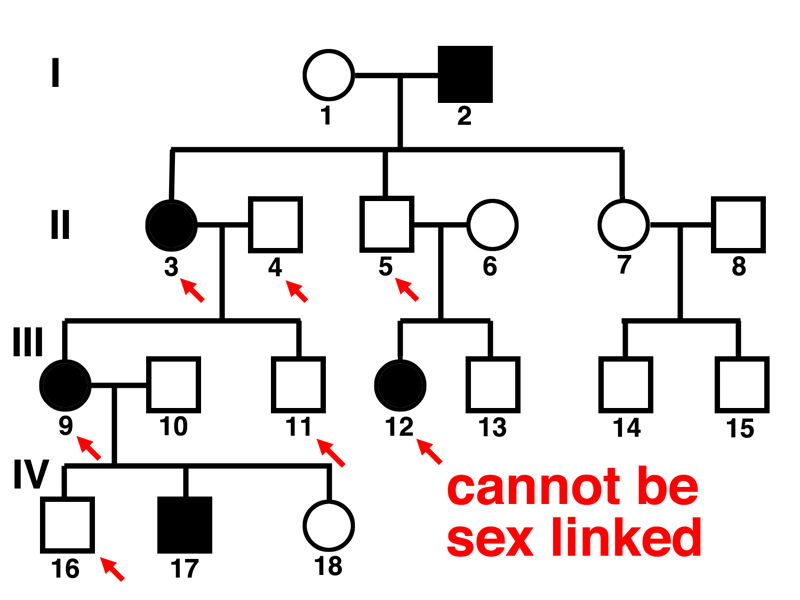
We can see in the pedigree that affected woman 3 has an unaffected son (11), affected woman 9 has an unaffected father (4), affected woman 9 has an unaffected son (16), and affected woman 12 has an unaffected father (5). We know that this is a recessive allele from the first question, so we have eliminated the hypothesis of a sex-linked recessive. This must be an autosomal recessive.
Some of the first evidence that suggested to us that genes are on chromosomes came from our consideration of human sex chromosome aneuploids resulting from sex chromosome nondisjuction, which we discussed in the second lecture.
In the Punnett square shown below, we consider the products of maternal nondisjunction of the X chromosome, fertilized by either X-bearing or Y-bearing sperm. The first column (normal meiosis to produce an X-bearing egg) is much more frequent than either of the exceptional types (XX and 0).
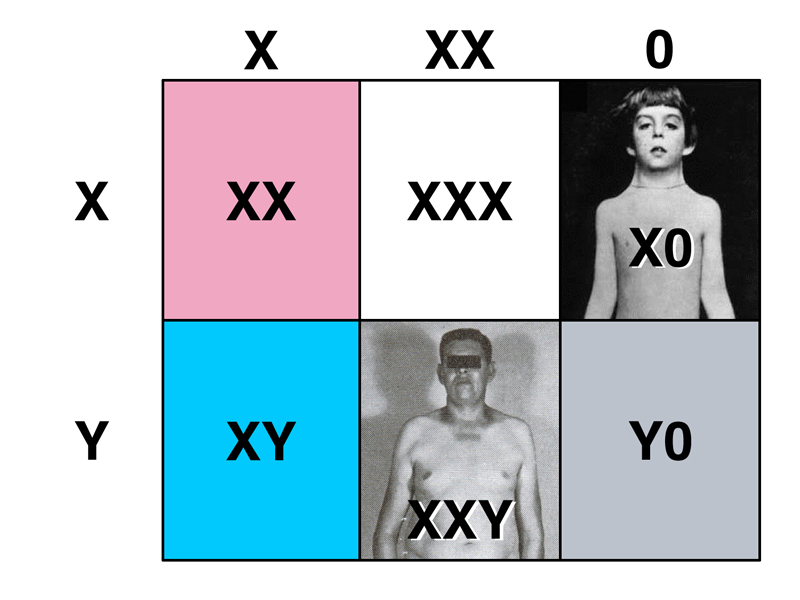
We know that XX individuals are normal females, XY individuals are normal males, Y0 individuals are inviable, X0 individuals are females with Turner Syndrome, and XXY individuals are males with Klinefelter Syndrome. We also learned that XXX individuals are completely normal females, who are not usually aware of their condition.
The study of human sex chromosome aneuploids suggests that genes are on chromosomes. We can see that the X chromosome is required for development, because YO individuals are inviable. The Y chromosome appears to determine maleness, because XXY individuals are male and X0 individuals are female.
The proof that genes are on chromosomes actually came from the study of the behavior of the white gene and other sex-linked genes in Drosophila, and was published by Bridges in 1916.
Returning to the crosses with white that we discussed earlier, recall that when we cross white-eyed females to red-eyed males, the offspring are all red-eyed females and white-eyed males.

This is only approximately correct. If we perform this experiment at sufficient scale, we will observe rare exceptional progeny at a frequency of around 1/2000: red-eyed males and white-eyed females, as shown below.
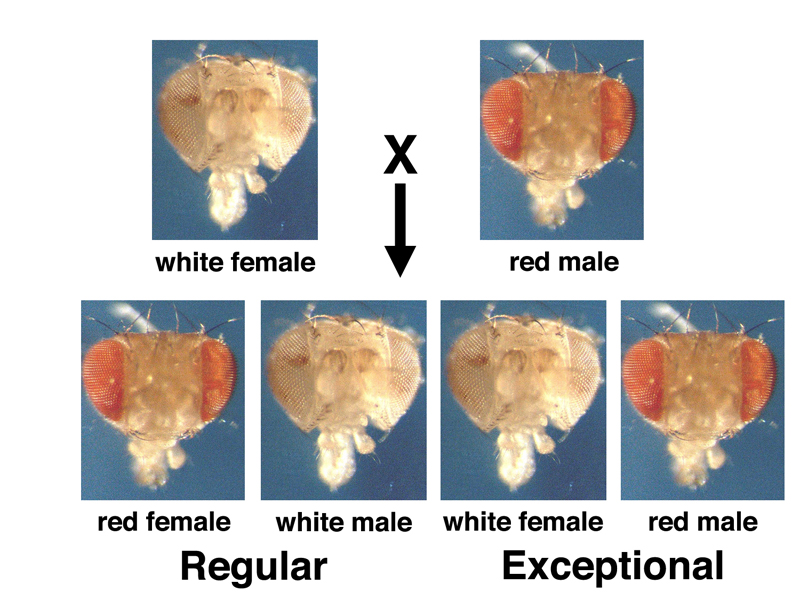
When Bridges tried to breed the exceptional red-eyed males, he found that they were invariably sterile. The exceptional white-eyed females were fertile and phenotypically normal in every way, except that when they were bred to red-eyed males, they gave a very high frequency of exceptions compared to their mothers. Exceptional progeny occurred at a frequency of around 2-3% from exceptional females.
The frequency at which exceptions occur in the first place (1/2000) rules out mutation as an explanation. Several students thought that sex chromosome nondisjunction might be responsible, but were troubled by what would happen if an egg with two X chromosomes (both bearing w) was fertilized by a Y-bearing sperm, because we have seen that XXY individuals are male in humans. One student suggested that such an egg could be fertilized by a sperm bearing neither an X nor a Y chromosome to produce an XX female homozygous for w.
This explanation requires nondisjunction in both the male and the female parent. As meiosis in the two parents is independent, the observed frequency of exceptional females would be the product of the frequency of sex chromosome nondisjunction in males and the frequency of sex chromosome nondisjunction in females. As the product is 1/2000, the frequency of nondisjunction in males and females (assuming an equal frequency) would be 1/44. It should be possible to detect this very high rate of sex chromosome nondisjunction in other experiments. As it happens, this is not the correct explanation.
A more direct experiment, which is what Bridges did, was to examine cells in mitosis from exceptional individuals to determine their karyotype. He found that exceptional females are XXY and exceptional males are X0.
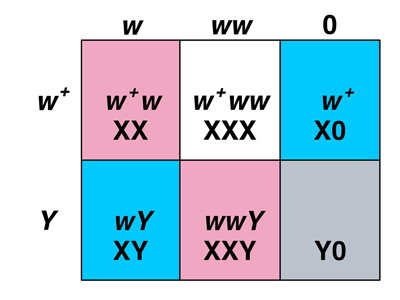
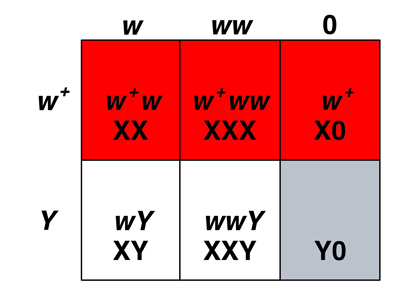
We can understand the origin of exceptional males and females by examining the Punnett squares below.
 |
 |
The Punnett square on the left shows us the sex of Drosophila sex chromosome aneuploids. We know that among the progeny from regular gametes, XX is female and XY is male. We know from the karyotype of exceptional males and females that XXY is female and X0 is male. Y0 is inviable. What about XXX? We know from other experiments that these are very poorly viable metafemales, which have an abnormal appearance.
The Punnett square on the right shows us that the white-eyed exceptional females are XXY. They have inherited two X chromosomes from their mothers and a Y chromosome from their fathers. Red-eyed exceptional males are X0. They have inherited an X chromosome from their fathers and no sex chromosome at all from their mothers.
We began a discussion of how the results of this experiment proves that genes are on chromosomes. A student pointed out that the sterility of X0 males shows that there is at least one factor required for male fertility on the Y chromosome, an excellent point.
Bridges argued that because in those rare cases where we see exceptional inheritance of sex-linked markers like white, the karyotype of those individuals is always consistent with the hypothesis that genes are on chromosomes. Exceptional females inheriting two w alleles from ther mothers always inherit two X chromosomes from their mothers and a Y chromosome from their fathers. Exceptional males inheriting the w+ allele from their fathers have inherited no sex chromosome from their mothers. The unusual karyotpes (XXY and X0) are not found among regular progeny, establishing that the abnormal inheritance of sex-linked alleles is always associated with an abnormal karyotype. Therefore, genes are on chromosomes.
This leads us to compare the rules for sex determination in Drosophila and humans.
| Drosophila female | Drosophila male | Human female | Human male |
| XX | XY | XX | XY |
| XXY | X0 (sterile) |
X0 (Turner) |
XXY (Klinefelter) |
| XXX (metafemale) |
XYY | XXX | XYY |
| In Drosophila, individuals with a single X are male. The Y chromosome is not involved in sex determination. |
In humans, individuals with at least one Y chromosome are male. The X chromosome is not involved in sex determination. |
||
A chromosomal sex determination system requires a dosage compensation system to adjust the level of gene expression in some way so that the ratio of gene expression from the X chromosome to gene expression from the autosomes is comparable in the two sexes. We had previously seen the effects of trisomy for various human chromosomes, together with a gene count for each chromosome, shown below.
| Chromosome | Gene Count | Trisomy |
| X | 1376 | normal |
| 12 | 1370 | inviable |
| 14 | 1315 | inviable |
| 16 | 1098 | inviable |
| 15 | 998 | inviable |
| 20 | 768 | inviable |
| 22 | 749 | inviable |
| 13 | 545 | Patau syndrome very poor survival |
| 18 | 429 | Edwards syndrome poor survival |
| 21 | 360 | Down Syndrome good survival |
| Y | 284 | ? |
Because XXX individuals (and even XXXX and XXXXX) are viable and indeed normal, even though the X chromosome has more genes than smaller chromosomes for which trisomy is not tolerated, we know that there must be a dosage compensation system in humans that makes our species relatively insensitive to the number of X chromosomes.
In contrast, in Drosophila, XXX metafemales are very poorly viable. We have already seen that Drosophila uses a different sex determination system. It is evident that it must have a different dosage compensation system as well.
We closed with a discussion of sex determination systems among animals. Some reptiles have a sex determination system based on temperature. Birds have a sex determination system in which females are heterogametic and males are homogametic. In honeybees, males are haploid, developing from unfertilized eggs. We concluded from this discussion that there is a great diversity of sex determination systems among animals, with many animals lacking sex chromosomes. Those animals that have a chromosomal sex determination system must also have a dosage compensation system; these are known to operate by a variety of different mechanisms in different animals.