| Home | Syllabus | Schedule | Lecture Notes | Extras | Glossary |
| Home | Syllabus | Schedule | Lecture Notes | Extras | Glossary |
We began by reminding everyone that Exam 3 is scheduled for Tuesday, November 12, covering all lecture material following Exam 2. We will hold a brief review of the material next lecture, and post the Study Guide by Thursday, November 7. Students would do well to review the material for Exam 2, as it is difficult to understand regulation without a solid grasp of transcription and translation.
We being our discussion of cancer genetics by looking at some fairly recent data on causes of death in the USA, shown below, which shows the age-adjusted death rate by cause.
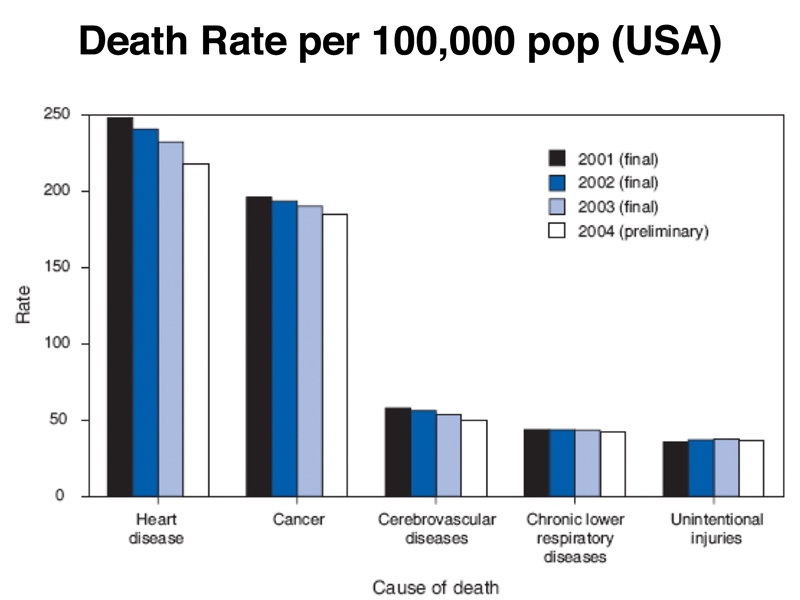
Heart disease is still the leading cause of death, followed closely by cancer. About 1/2 of men will get cancer in their lifetimes, and about 1/3 of women, although many who get cancer will not die of it. A show of hands revealed that most students know someone who had cancer.
The table below shows cancer incidence by type, reporting the estimate of new cases that will be seen by type in 2013, the number of deaths by type in 2013, the expected 5-year survival and the percent of all cancer deaths for each type. Only the most common cancer types are shown (there are hundreds of different kinds of cancer).
| Cancer Type | New cases | Deaths | 5 year survival | % cancer deaths |
| prostate | 238,590 | 29,720 | 99.2% | 5.1% |
| breast | 232,340 | 39,620 | 89.2% | 6.9% |
| lung | 228,190 | 159,480 | 16.6% | 27.5% |
| colorectal | 142,820 | 50,830 | 64.9% | 8.8% |
| melanoma | 76,690 | 9,480 | 91.3% | 1.6% |
| bladder | 72,570 | 15,210 | 77.9% | 2.6% |
| non-Hodgkin lymphoma | 69,740 | 19,020 | 69.0% | 3.3% |
We can see several things from the data. First of all, adding up just the cancers shown, there will be over a million new cases of cancer reported in 2013. There are about 330 million people in the United States, so 0.3% of the population of the United States will learn that they have cancer in 2013.
The five-year survival for the different cancer types varies widely. While 99.2% of men diagnosed with prostate cancer will be alive after five years, only 16.6% of patients with lung cancer will survive five years. Lung cancer accounts for over a quarter of all cancer deaths. This tells us that diagnosis and treatment for different types of cancer is very different, and suggests that it is misleading to speak of cancer as a single disease. It is more useful to think of cancer as a collection of related diseases.
There is a strong genetic component to cancer. There are fifty or so hereditary cancer predispositions, in which people inherit a greatly elevated risk of a specific type of cancer. The most common of these hereditary cancer predispositions is Hereditary Breast and Ovarian Cancer. Risk alleles of two genes, BRCA1 and BRCA2, confer a greatly elevated risk of breast and ovarian cancer. A pedigree in which a BRCA1 risk allele is segregating is shown below.
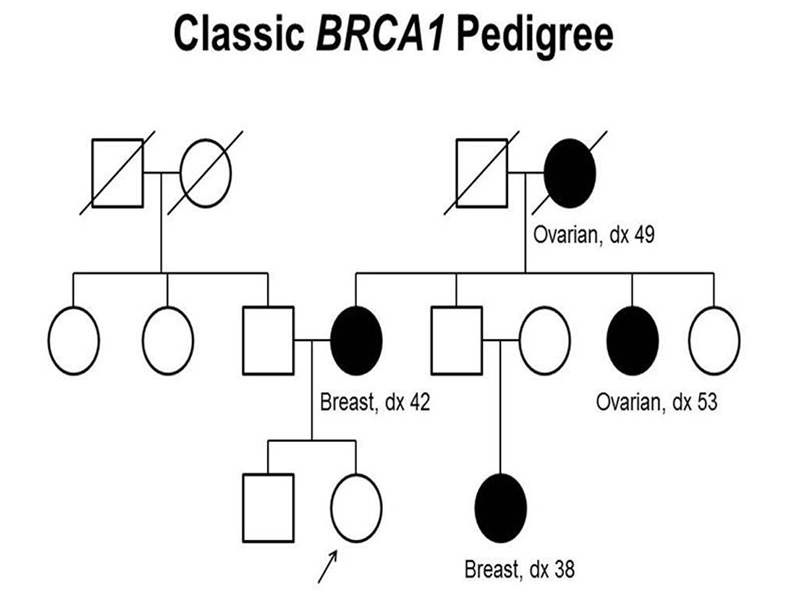
We see in the pedigree that the trait behaves as a dominant mutation. Notice that the affected female in the third generation has an unaffected father who is an obligate carrier. Although men who carry a risk allele for BRCA1 are at greatly elevated risk for breast cancer, the incidence in men is still very low, as shown below.
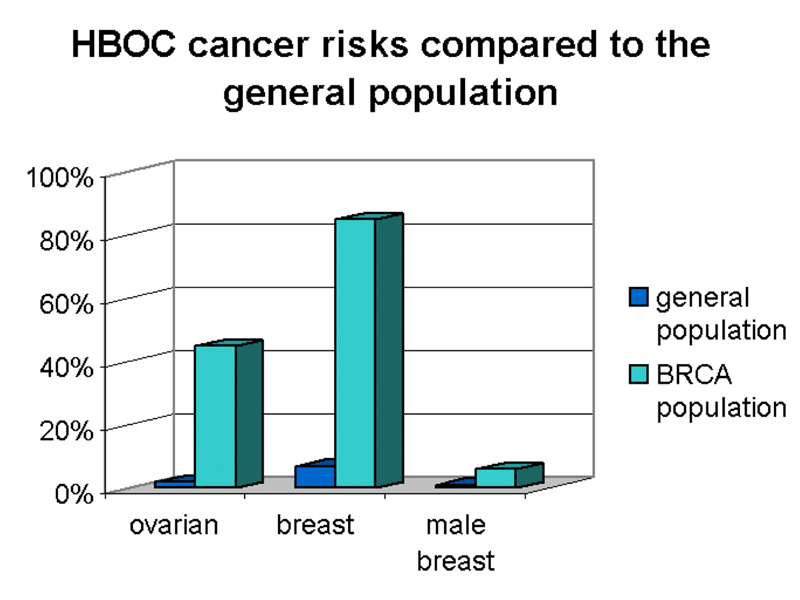
As shown in the figure, women who carry a risk allele of BRCA1 have about a 40% risk of developing ovarian cancer and an 80% risk of developing breast cancer. Men who carry an risk allele of BRCA1 have a slight risk of developing breast cancer.
All people carrying risk alleles of BRCA1 are heterozygotes. In kindreds where there are risk alleles on both sides of the family, compound heterozygotes are never seen, suggesting that complete loss of function of BRCA1 is inconsistent with viablity. Studies in the mouse support this. When mice heterozygous for loss-of-function alleles are bred, the homozygotes die as embryos.
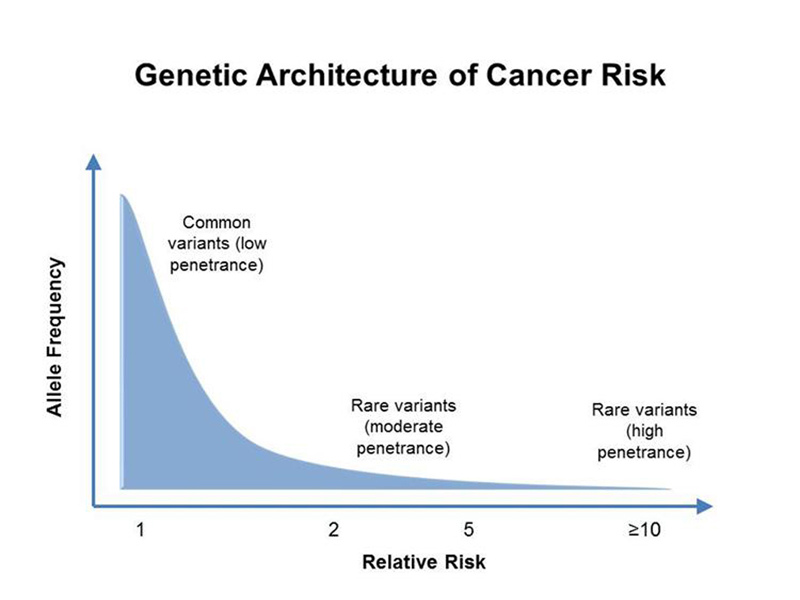
In general, as shown below, cancer risk alleles with high penetrance have a low allele frequency in human populations due to selection. Moderate risk alleles have a higher allele frequency, and there are a large number of common genetic variants with high allele frequencies that slightly elevate cancer risk. It is easier to analyze risk alleles with high penetrance, so our view of hereditary cancer predisposition is somewhat distorted.
It is clear that the average person's risk of developing cancer is strongly correlated with environmental factors. When asked for an environmental factor that increased cancer risk, a large number of students offered cigarette smoking and lung cancer as an example, showing that the millions spent on public service announcements have not been wasted. Other examples included oral cancer and smokeless tobacco (or betel nut outside the USA), lung cancer and radon exposure, and cancer clusters in communities located on Superfund sites, where toxic chemicals have been dumped.
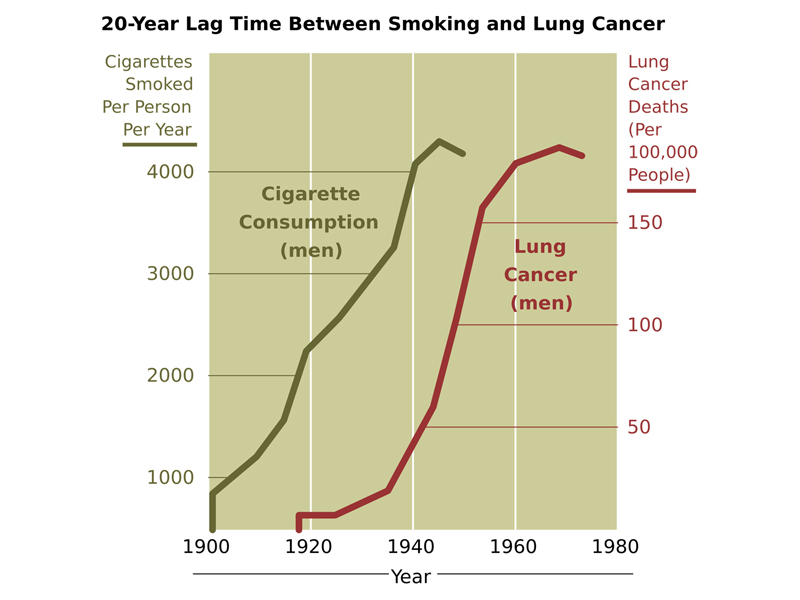
The figure below shows the 20-year time lag between the start of the mass marketing of cigarettes and an increase in the number of cases of lung cancer in men. Because it was initially not socially acceptable for women to smoke, the rise in the incidence of smoking-related lung cancer in women occurs later (not shown).
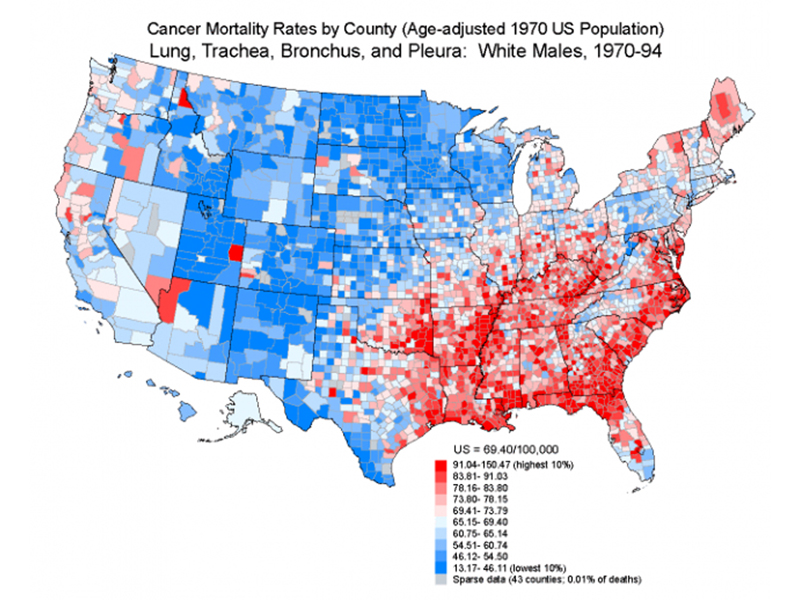
The figure below shows the incidence of lung and related cancers among white men by county for the time period of 1970 to 1994. The map somewhat resembles the obesity map that we saw in an earlier lecture. We speculated on the meaning of the high incidence of lung cancer in the Southeast and along the Mississippi River, noting that most of the highest risk counties either grow tobacco or are located near tobacco-growing areas. It is also likely that smoking is inversely correlated with level of education and socioeconomic status. In any case, the map certainly does not reflect a geographic difference in the frequency of risk alleles for cancer, so whatever the cause, it is environmental.
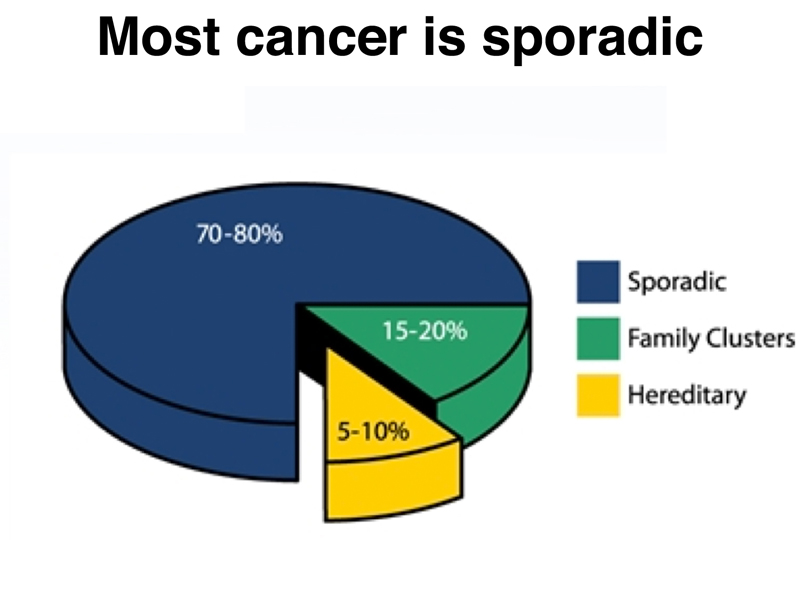
Early in the course, we emphasized that most genes interact with other genes and with the environment. If we consider cancer as a single disease, which it is not, we can quantify this interplay as shown below. Most cancer is sporadic, meaning that it is not associated with the inheritance of risk alleles and that it does not cluster in families. About 5-10% of cases of cancer are hereditary, resulting from the inheritance of risk alleles that greatly increase a person's chance of developing cancer. About 15-20% of cases of cancer occur in family clusters. This can be the result of enviromental factors, such as shared exposure to carcinogens because of the location of the home, or perhaps the result of genetic factors, such as risk alleles with low penetrance.
We began our discussion of the biology of cancer with a simple description of the disease. Cancer is a disease in which normal control of cell division is lost in some cells. The current model of cancer proposes that somatic mutation is the cause of a cell's alteration in the normal control of cell division. This model is supported by a large body of work that demonstrates that every chemical that can be shown to be carcinogenic is also a mutagen. Multiple mutations are required to transform a normal cell into a cancer cell.
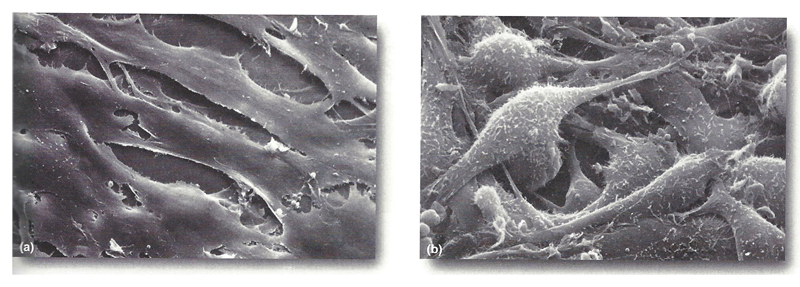
Some aspects of the phenotype of cancer cells are evident in tissue culture. The figure below shows normal cultured cells on the left and cancer cells on the right. The appearance of the cancer cells is altered. They are "hairy," with many projections of the plasma membrane. The normal cells show contact inhibition in culture, that is, they spread out and don't grow on top of each other. The cancer cells do not show contact inhibition. It is evident that the culture is several cell layers thick.
Progression to a malignant tumor occurs in stages, with visible changes at each stage evident in some types of cancer. The figure below shows the progression to malignant colon cancer.
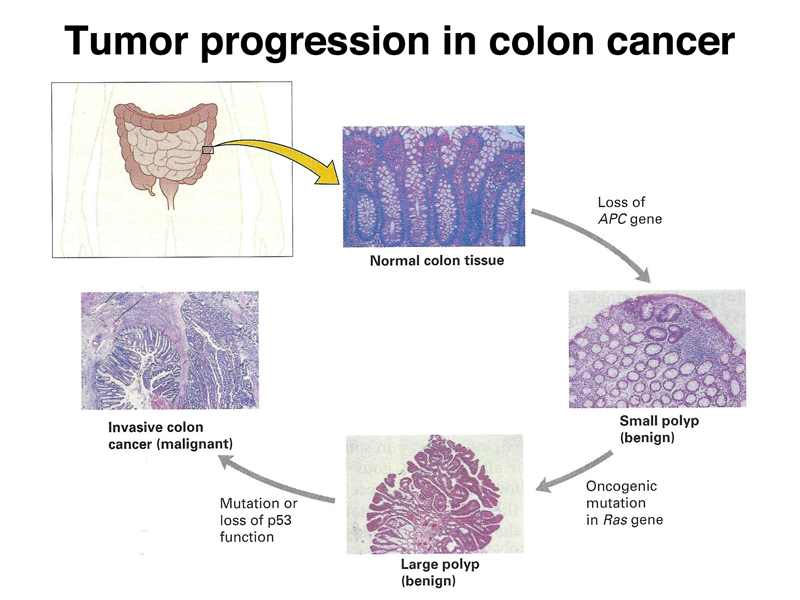
The first step is the occurrence of small benign polyps. This requires the loss of the APC protein, discussed below. Progression to a large benign polyp requires a mutation in the Ras oncogene, discussed below. The progression to malignancy requires the loss of function of Tp53, also discussed below. Different types of cancer require the loss of function of different specific genes, although mutations in some specific genes are found in many different types of cancer. All cancers require multiple somatic mutations.
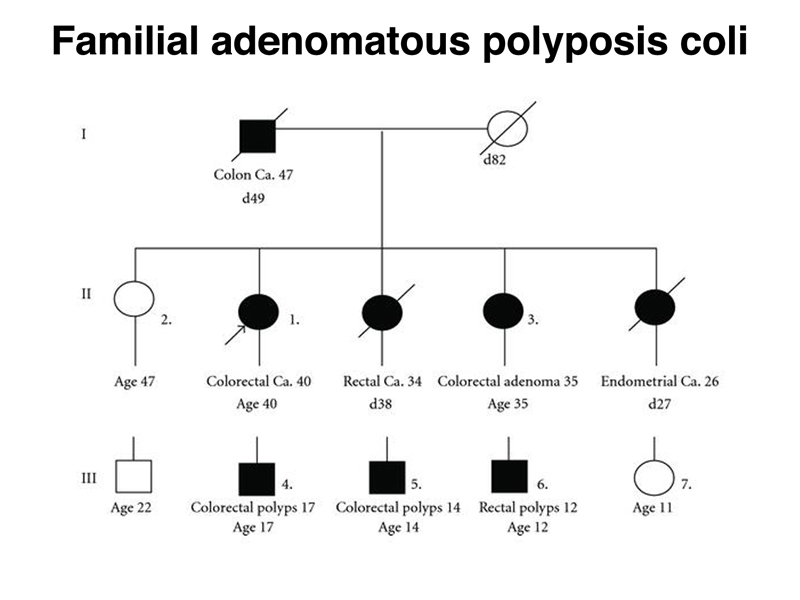
The figure below shows a pedigree for Familial adenomatous polyposis coli (FAP). Note that there are multiple affected individuals in three generations. In the first two generations, there are mutliple instances of cancer with an early age of onset. In the third generation, there are three affected individuals with benign polyps in their teenage years. FAP is due to inheritance of defects in the APC gene. The figure above shows that mutations in this gene are required for the occurrence of benign polyps. The pedigree shows that FAP is a dominant condition. Inheritance of a loss-of-function allele of APC confers FAP.
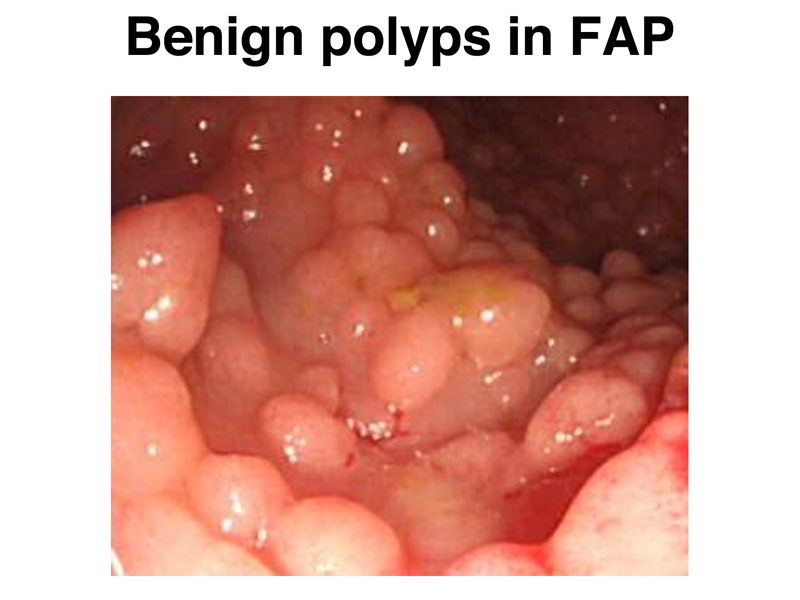
The image below shows multiple polyps in the colon of someone with FAP. It is not uncommon for these patients to have hundreds or even thousands of polyps.
The role of the APC gene product in signal transduction is now known. As shown in the figure below, APC is a protein in the Wnt signalling pathway. Together with β-catenin, it acts to negatively regulate genes that promote cell division. In the absence of APC, cell division is partially deregulated.
Retinoblastoma is a tumor of the retina that occurs in children. Familial retinoblastoma is inherited as a dominant condition. In familial retinoblastoma, both eyes are affected, and there are multiple tumors in each eye. In sporadic retinoblastoma, there is no family history, and only one eye is affected. The figure below shows the retinas of an individual with familial retinoblastoma before and after treatment with chemotherapy and radiation.
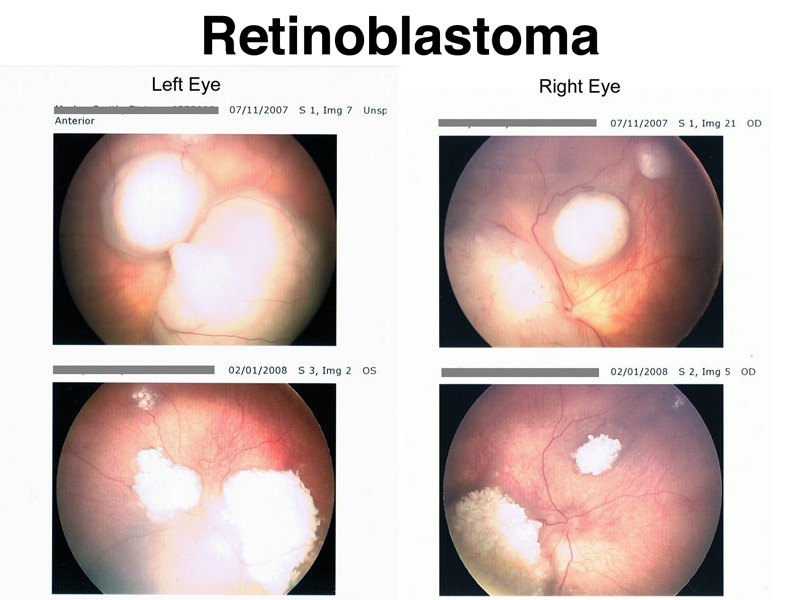
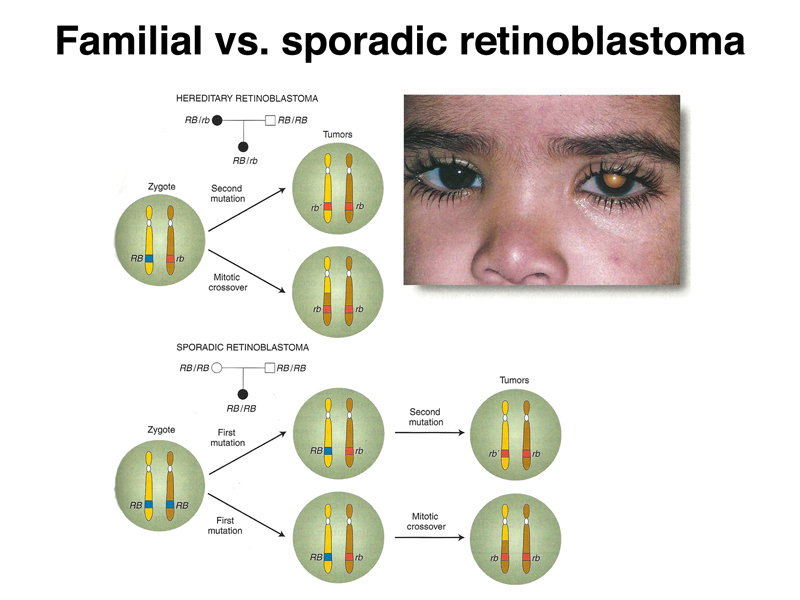
People with familial retinoblastoma have inherited a loss-of-function allele of the RB1 gene. Although this greatly increases their risk for retinoblastoma, retinoblastoma only arises in cells that have no functional copy of the RB1 gene. As shown below, the tumors arising in these patients occur because retinal cells have lost the other copy by point mutation, mitotic recombination, or some other event that causes loss of heterozygosity. In sporadic retinoblastoma, cells that become tumors must first have a somatic mutation in RB1, followed by losing the other copy of RB1 by point mutation, mitotic recombination, or some other event that causes loss of heterozygosity.
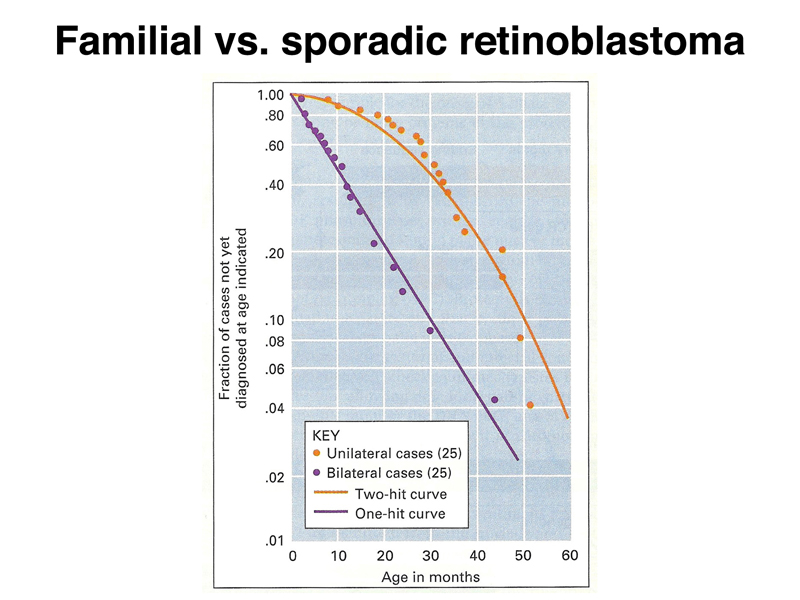
Dividing retinoblastoma cases into bilateral (familial) and unilateral (sporadic) cases reveals that the familial cases have an earlier age of onset, and the age of occurrence follows an exponential plot, as shown in the semilog plot below. In contrast, the age of occurrence in sporadic cases plots differently. Familial retinoblastoma occurs with "one-hit" kinetics, while sporadic retinoblastoma occurs with "two-hit" kinetics. The same is true for the other cases of heritable cancer predisposition that we have discussed, breast and ovarian cancer in individuals heterozygous for loss-of-function alleles of BRCA1 or BRCA2, and colon cancer in individuals heterozygous for loss-of-function alleles of APC.
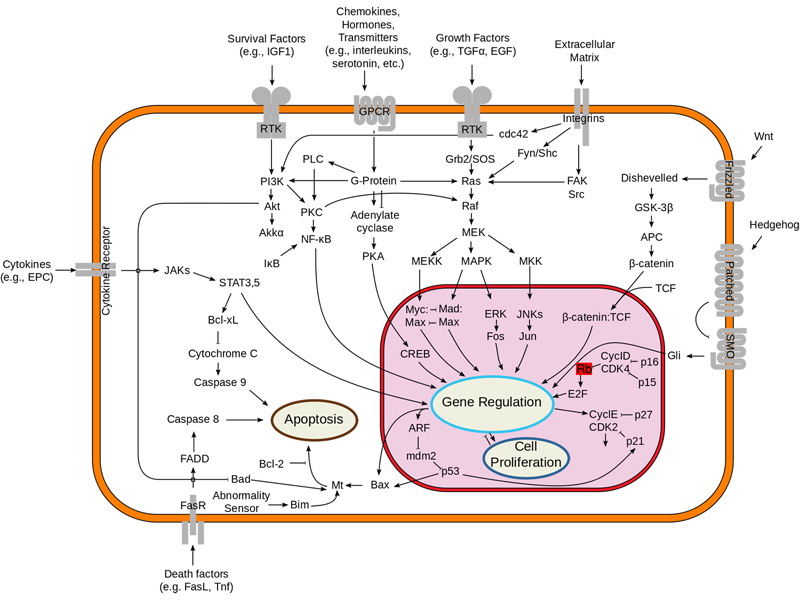
The figure below shows where the RB1 gene product fits in an overview of signal transduction. Like APC, the gene product of RB1 negatively regulates genes required for cell division.
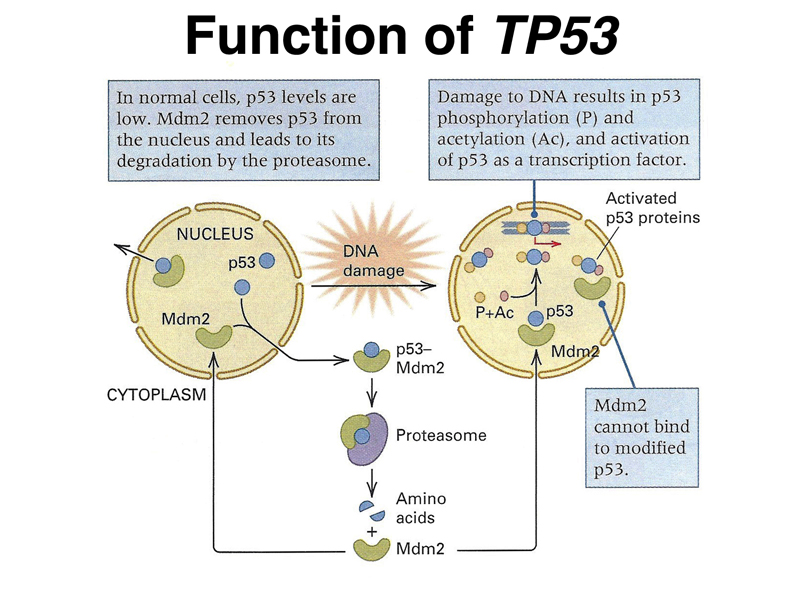
The last example of this kind that we will consider is Li-Fraumeni Syndrome, an inherited cancer predisposition that results in an increased risk for a wide variety of tumors. Li-Fraumeni Syndrome is inherited as a dominant condition resulting from heterozygosity for a loss-of-function allele of TP53. As shown below, the product of the TP53 gene (p53) is part of a system of responding to DNA damage. Under normal conditions, the p53 protein binds to another protein called Mdm2 and is destroyed by the proteasome. In the presence of DNA damage, p53 is post-translationally modified by phosphorylation and acetylation. This makes p53 unable to bind to Mdm2, and p53 is activated as a transcription factor.
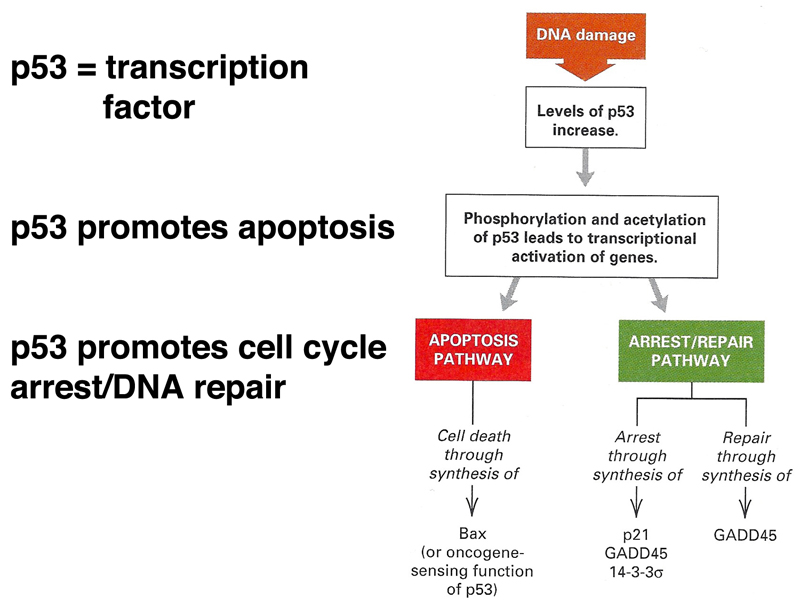
The consequences of activation of p53 are shown in the figure below. Activated p53 promotes apoptosis (programmed cell death), cell cycle arrest, and DNA repair. This gives the cell a chance to attempt DNA repair before undertaking cell division. If that is not successful, the cell undergoes programmed cell death.
The figure below shows where p53 fits in an overview of signal transduction.
All of the genes that we have discussed so far (BRCA1, BRCA2, APC, RB1, and TP53) are classified as tumor-suppressor genes. Being heterozygous for a loss-of-function allele greatly increases an individual's risk for specific cancers. The tumor cells themselves are homozygous or hemizygous for loss-of-function alleles of these genes, so at the cellular level, tumor-suppressor genes are recessive.
There is another class of genes involved in cancer, called oncogenes. One of the first oncogenes discovered came from the study of Rous sarcoma virus, a retrovirus (RNA virus) that causes sarcomas in chickens. Students were able to name a couple of human cancer viruses, specifically human papilloma virus, which causes cervical cancer, and hepatitis virus, which causes liver cancer. Both HPV and hepatitis are carcinogenic because they integrate into many different positions in the genome, acting as mutagens. The mechanism by which Rous sarcoma virus causes cancer in chickens is different, as shown in the figure below.
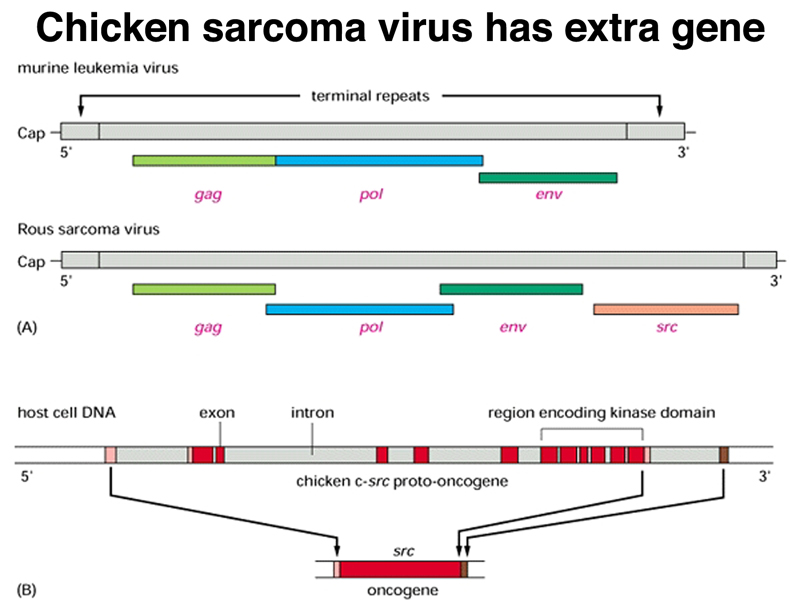
Compared to closely related retroviruses like murine leukemia virus, there is an extra gene in Rous sarcoma virus, named src. The src gene carried by Rous sarcoma virus causes tumors; if it is inactivated, the virus does not induce sarcomas. Surprisingly, the src gene carried by the virus (v-src) is a mutated version of a gene found in normal cells (c-src). The protein encoded by the src gene is a tyrosine kinase. The v-src gene has a gain-of-function mutation that makes the kinase constitutively activated, not subject to the regulation that controls the c-src gene.
The position of src in the signal transduction overview is shown in the figure below.
There is another oncogene called v-erbB, identified in a different tumor virus, that encodes an activated form of the receptor for epidermal growth factor (EGF). The normal cellular gene, c-erbB, encodes a receptor tyrosine kinase, which when bound to epidermal growth factor, sends a signal via its cytoplasmic tyrosine kinase domain that promotes cell division. The oncogenic form encoded by the virus is partially deleted at the amino terminus and the carboxyl terminus, and encodes a constitutively-activated form of the receptor that sends a mitogenic signal in the absence of EGF. Like v-src, the constitutively-activated v-erbB gene product promotes cell division even in the presence of the normal form of the gene, c-erbB.
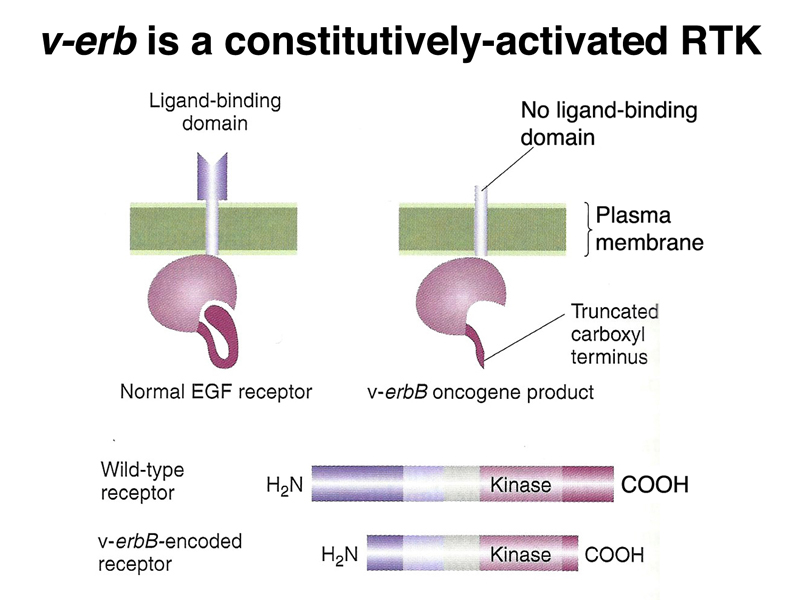
The position of the gain-of-function allele v-erbB in the signal transduction overview is shown below. The EGF receptor is one of many receptor tyrosine kinases.
Oncogenic mutations in RAS are common in many different cancers. As shown below, a single amino acid substitution in the RAS gene can eliminate the ability of the protein to hydrolyze GTP to GDP. The mutant form of the protein will therefore always be bound to GTP, and hence will be continuously active. Activated RAS will send signals to stimulate cell division even in the presence of the normal protein encoded by the other allele.
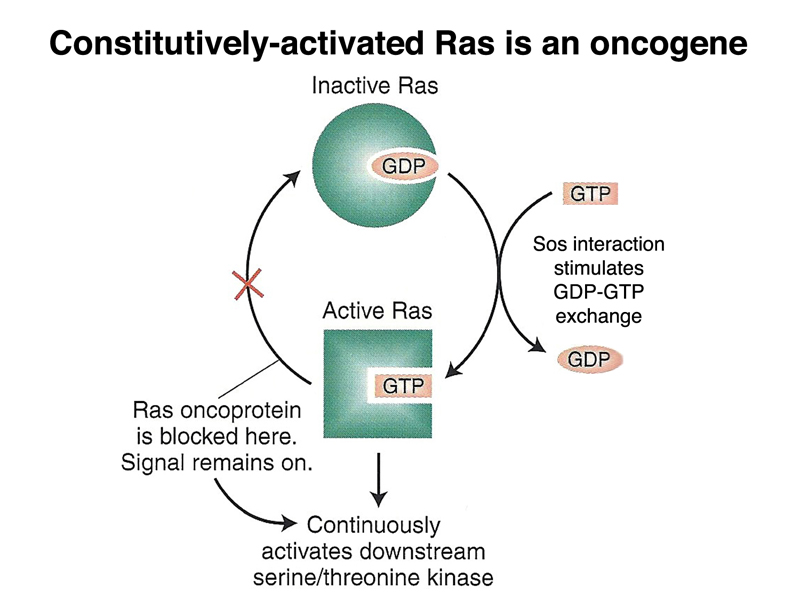
The central position of RAS in signal transduction is shown below. RAS signalling is downstream of all receptor tyrosine kinases and G-protein coupled receptors.
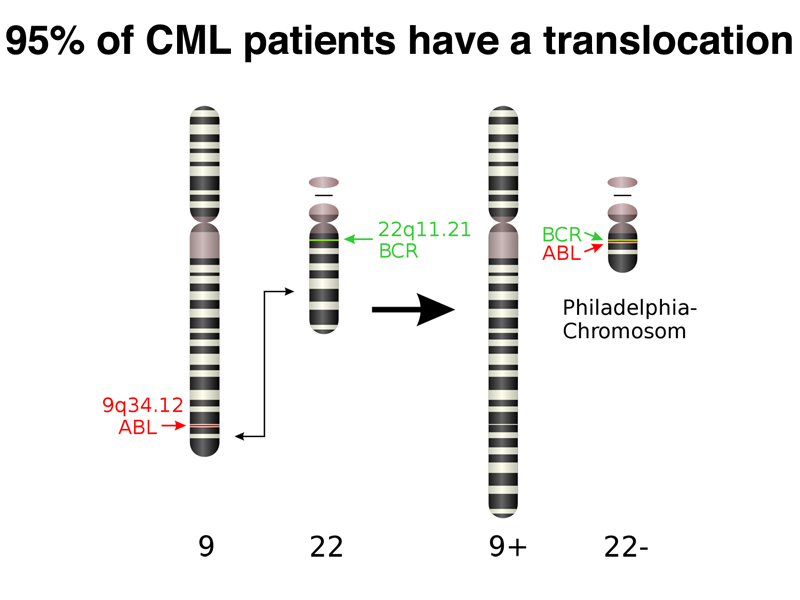
There is an unusual oncogenic mutation that is common in Chronic Myelogenous Leukemia (CML): 95% of CML patients have a specific reciprocal translocation, called the Philadelphia chromosome, in their tumor cells. This chromosome rearrangement, shown below, occurs somatically.
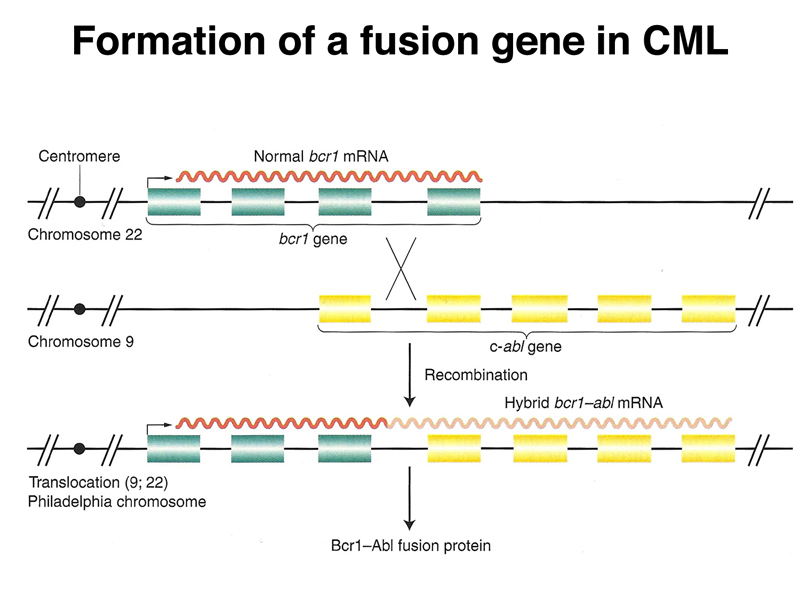
There is no change in ploidy associated with a reciprocal translocation, so there must be something oncogenic about the rearrangement breakpoint. This is shown below. All occurrences of the Philadelphia chromosome in CML have breakpoints somewhere in a gene called bcr1 (for breakpoint cluster 1) that creates a gene fusion with the ABL gene. The ABL gene is a receptor tyrosine kinase. The fusion gene expresses a constitutively-activated receptor tyrosine kinase like the v-erbB oncogene.
The position of the activated ABL receptor tyrosine kinase in the overview of signal transduction is shown below.
Another example of an oncogenic chromosome rearrangement is shown below. In this case, a reciprocal translocation produces lymphomas. The translocation creates a fusion gene that expresses BCL-2 in lymphocytes.
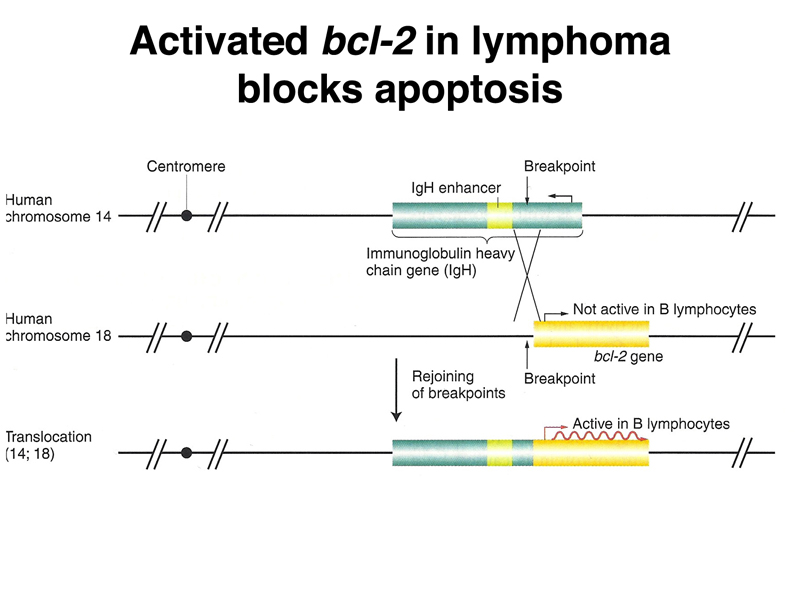
The BCL-2 protein acts together with the BAX protein to balance the cell between immortalization and programmed cell death. In the presence of excess BAX, apoptosis occurs. In the presence of excess BCL-2, the cell is immortalized. In normal cells, the two gene products are in balance, and BAX/BCL-2 heterodimers form. This prevents BAX/BAX homodimers, which promote apoptosis, and activation of BCL-2, which promotes immortality.
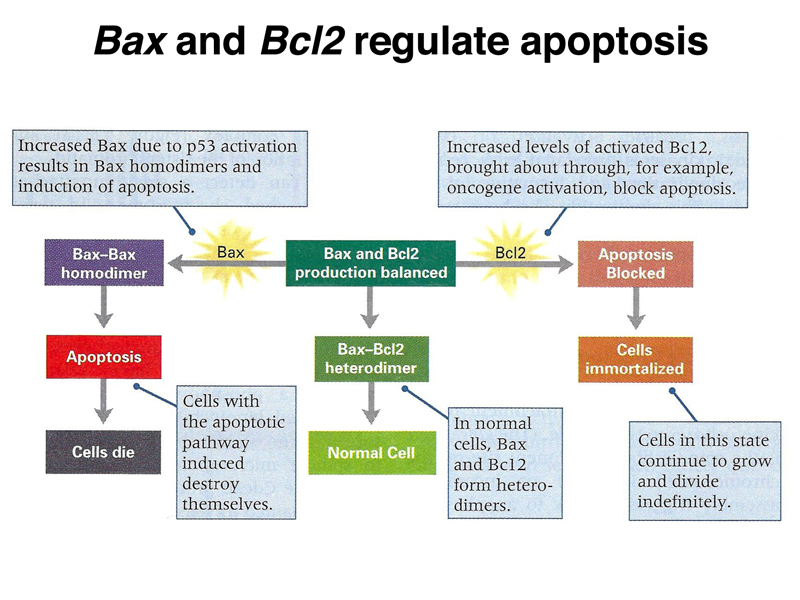
The position of BCL-2 in the signal transduction overview is shown below.
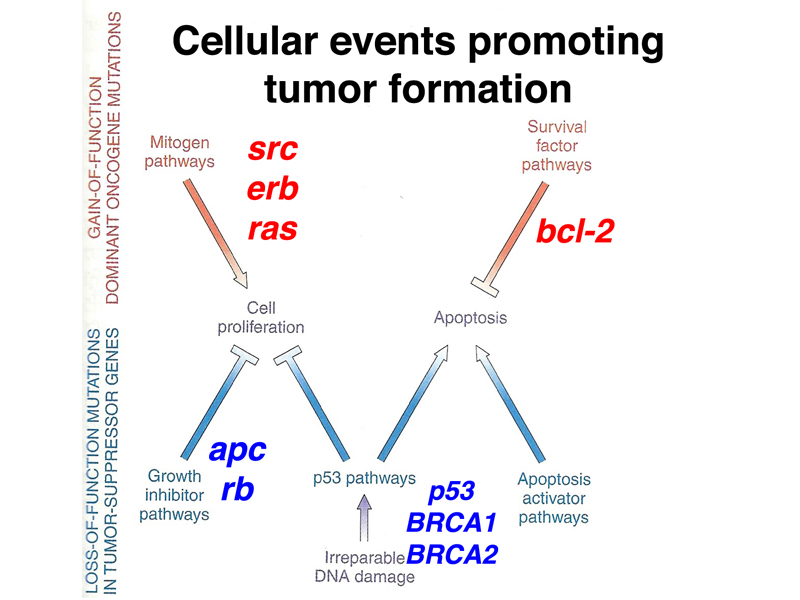
Cancer is a genetic disease that results from multiple somatic mutations in genes important for regulating cell division. We have discussed two kinds of mutations that are important in the development of cancer: loss-of-function alleles of a class of genes called tumor suppressors, and gain-of-function alleles of a class of genes called oncogenes. Gain-of-function alleles of oncogenes are dominant at the cellular level, acting to promote tumor formation even in the presence of a normal allele. Loss-of-function alleles of tumor-suppressor genes are recessive at the cellular level, even though they act as dominant cancer risk factors in pedigrees.
The figure below shows the roles of the genes that we have discussed in stimulating or inhibiting cell proliferation or apoptosis. There are hundreds of genes important for the development of tumors of one type or another; we have reviewed some of the more important genes that are mutated in many tumor types (p53 and RAS) or some of the very well-studied examples (BRCA1, BRCA2, and RB1).